4 механизм инактивации g белков. В. структурно-функциональная организация g-белков. Другие белки, связывающие иммуноглобулины
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим образование вторичных посредников гормонального сигнала. G-белки - олигомеры, состоящие из α, β и γ-субъединиц. Состав димеров βγ незначительно различаются в разных тканях, но в пределах одной клетки все G-белки, как правило, имеют одинаковый комплект βγ-субъединиц. Поэтому G-белки принято различать по их α-субъединицам.. Выявлено 16 генов, кодирующих различные α-субъединицы G-белков. Некоторые из генов имеют более одного белка, вследствие альтернативного сплайсинга РНК.
Каждая α-субъединица в составе G -белка имеет специфические центры:
Связывания ГТФ или ГДФ;
Взаимодействия с рецептором;
Связывания с βγ-субъединицами;
Фосфорилирования под действием протеинкиназы С;
Взаимодействия с ферментом аденилатциклазой или фосфолипазой С.
В структуре G -белков отсутствуют α-спиральные, пронизывающие мембрану домены. G -белки относят к группе «заякоренных» белков (рис. 5-34).
Рис. 5-34. Положение G-белков в мембране. Для ассоциации G-белков важно ацилирование α-протомеров алифатическими радикалами жирных кислот, миристиновой кислоты (М) или изопреновой. γ-Субъединица G-белка имеет геранил-геранильную группу (Г), связанную тиоэфирной связью с остатком цистеина С-конца.
Регуляция активности G -белков
Различают неактивную форму G -белка - комплекс αβγ-ГДФ и активированную форму αβγ -ГТФ. Активация G-белка происходит при взаимодействии с комплексом активатор-рецептор, изменение конформации G-белка снижает сродство α-субъединицы к молекуле ГДФ и увеличивает к ГТФ. Замена ГДФ на ГТФ в активном центре G-белка нарушает комплементарность между α-ГТФ и βγ-субъединицами. Рецептор, связанный с сигнальной молекулой, может активировать большое количество молекул G-белка, таким образом обеспечивая усиление внеклеточного сигнала на этом этапе (рис. 5-35).

Рис. 5-35. Цикл функционирования G-белка. R s - рецептор; Г - гормон; АЦ - аденилатциклаза.
Активированная α-субъединица G-белка (α-ГТФ) взаимодействует со специфическим белком клеточной мембраны и изменяет его активность. Такими белками могут быть ферменты аденилатциклаза, фосфолипаза С, фосфодиэстераза цГМФ, Na + -каналы, К + -каналы.
Следующий этап цикла функционирования G-белка - дефосфорилирование ГТФ, связанного с α-субъединицей, причём фермент, катализирующий эту реакцию, - сама α-субъединица.
Дефосфорилирование приводит к образованию комплекса α-ГДФ, который не комплементарен специфическому белку мембраны (например, аденилатциклазе), но имеет высокое сродство к βγ-протомерам. G-белок возвращается к неактивной форме - αβγ-ГДФ. При последующей активации рецептора и замене молекулы ГДФ на ГТФ цикл повторяется снова. Таким образом, α-субъединицы G-белков совершают челночное движение, перенося стимулирующий или ингибирующий сигнал от рецептора, который активирован первичным посредником (например, гормоном), на фермент, катализирующий образование вторичного посредника.
Некоторые формы протеинкиназ могут фосфорилировать α-субъединицы G-белков. Фосфорилированная α-субъединица не комплементарна специфическому белку мембраны, например, аденилатциклазе или фосфолипазе С, поэтому не может участвовать в передаче сигнала.
Г. аденилатциклаза
Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ (рис. 5-36), - ключевой фермент аденилатциклазной системы передачи сигнала. Аденилатциклаза обнаружена во всех типах клеток.

Рис. 5-36. Образование циклического аденозинмонофосфата (цАМФ).
Фермент относят к группе интегральных белков клеточной мембраны, он имеет 12 трансмембранных доменов. Внеклеточные фрагменты аденилатциклазы гликозилированы. Цитоплазматические домены аденилатциклазы имеют два каталитических центра, ответственных за образование цАМФ - вторичного по средника, участвующего в регуляции активности фермента протеинкиназы А.
На активность аденилатциклазы оказывают влияние как внеклеточные, так и внутриклеточные регуляторы. Внеклеточные регулятор (гормоны, эйкозаноиды, биогенные амины) осуществляют регуляцию через специфические рецепторы, которые с помощью α-субъединиц G-белков передают сигналы на аденилатциклазу. αs - Субъединица (стимулирующая) при взаимодействии с аденилатциклазой активирует фермент, αi- Субъединица (ингибирующая) ингибирует фермент. В свою очередь, аденилатциклаза стимурирует проявление ГТФ- фосфотазной активности α- субъединиц. В результате дефосфорилирования ГТФ образуются субъединицы а s -ГДФ и а i -ГДФ, не комплементарные аденилатциклазе.
Из 8 изученных изоформ аденилатциклазы 4 - Са 2+ -зависимые (активируются Са 2+). Регуляция аденилатциклазы внутриклеточным кальцием позволяет клетке интегрировать активность двух основных вторичных посредников цАМФ и Са 2+ .
Д. фосфолипазы
Фосфолипазы - ферменты класса гидролаз, катализирующие катаболизм глицерофосфолипидов. Различают фосфолипазы секреторные, входящие в состав панкреатического сока, и клеточные фосфолипазы. Клеточные фосфолипазы А 1 , А 2 , D, С различаются по специфичности к отщепляемой группе. Все фосфолипазы - кальций зависимые ферменты (рис. 5-37).

Рис. 5-37. Действие фосфолипаз.
Фосфолипаза С - фермент, гидролизующий фосфоэфирную связь в глицерофосфолипидах. В клетках человека идентифицировано 10 изоформ фосфолипазы С, различающихся по молекулярной массе, локализации, способу регуляции, субстратной специфичности. В структуре всех изоформ фосфолипазы С отсутствуют гидрофобные домены, которые могли бы обеспечить их взаимодействие с мембраной. Однако некоторые формы фосфолипазы С связаны с помощью гидрофобного «якоря» - ацильного остатка миристиновой кислоты или за счёт взаимодействия с поверхностью бислоя. Каталитическая активность всех изоформ фосфолипазы С зависит от ионов кальция.
Большинство фосфолипаз С специфично в отношении фосфатидилинозитолов и практически не гидролизует другие типы фосфолипидов. Активный фермент может гидролизовать до 50% от общего количества фосфатидилинозитолов клеточной мембраны. При гидролизе фосфатидилинозитол-4,5-бисфосфата (ФИФ 2) образуются продукты диацилглицерол (ДАГ) и инозитол-1,4,5-трифосфат (ИФ 3), служащие вторичными посредниками в трансмембранной передаче сигнала по инозитолфосфатному пути.
Е. протеинкиназы
Все полярные сигнальные молекулы, действующие на клетку-мишень через мембранные рецепторы, осуществляют свою биологическую функцию путём фосфорилирования специфических белков и ферментов, регулирующих метаболизм в клетке. Фосфорилирование изменяет (увеличивает или уменьшает) их активность. Катализируют фосфорилирование белков (протеинов) протеинкиназы по аминокислотным остаткам серина, треонина, тирозина. Протеинкиназы могут быть субъединицей мембранного рецептора, например тирозиновая протеинкиназа рецептора инсулина, активность которой регулируется гормоном. Другая группа - протеинкиназы, регулируемые вторичными вестниками гормонального сигнала (цАМФ, цГМФ, Са 2+ , ДАГ), например протеинкиназа А, протеинкиназа С, протеинкиназа G, кальмодулинзависимые протеинкиназы и др.
Протеинкиназы А
Протеинкиназы А (цАМФ-стимулируемые) участвуют в аденилатциклазной системе передачи сигнала. Протеинкиназа А состоит из 4 субъединиц R 2 С 2 - двух регуляторных субъединиц (R 2) и двух каталитических (С 2) (см. рис. 5-41). Комплекс R 2 С 2 не обладает ферментативной активностью.
Комплекс R 2 С 2 разными способами прикрепляется к мембране. Некоторые формы протеинкиназы А «заякориваются» с помощью алифатического остатка миристиновой кислоты каталитических субъединиц. Во многих тканях протеинкиназа А связана с «заякоренным» белком АКАР s (от англ. сАМР dependent protein kinase anchoring proteins). АКАР s имеет центр связывания для регуляторных субъединиц протеинкиназы А. С помощью белка АКАР s протеинкиназа А связывается с мембраной в области локализации ферментов, катализирующих образование цАМФ (аденилатциклаза) или его гидролиз (фосфодиэстераза), а также белков, в регуляции активности которых фермент принимает участие, например потенциалзависимые Са 2+ -каналы.
Регуляторные субъединицы протеинкиназы А имеют специфические центры для связывания цАМФ. Присоединение цАМФ к регуляторным, субъединицам приводит к изменению конформации последних и снижению сродства к каталитическим субъединицам С, происходит диссоциация по схеме:
цАМФ 4 + R 2 С 2 -> цАМФ 4 R 2 + С + С
Субъединицы С представляют собой активную форму протеинкиназы А, которая катализирует реакции фосфорилирования по серину и треонину. Каталитические субъединицы С у разных типов протеинкиназ а не идентичны, они различаются прежде всего специфичностью в отношении белков-субстратов.
Протеинкиназы С
Протеинкиназы С участвуют в инозитолфосфатной системе передачи сигнала. Фермент состоит из двух функционально различных доменов - регуляторного и каталитического. Регуляторный домен содержит 2 структуры («цинковые пальцы»), образованные фрагментами пептидной цепи, богатыми цистеином, и содержащими 2 иона цинка (см. раздел 1). «Цинковые пальцы» участвуют в связывании диацилглицерола. Другой фрагмент регуляторного домена имеет высокое сродство к Са 2+ . Повышение концентрации кальция в цитозоле увеличивает сродство протеинкиназы С к фосфатидилсерину мембраны. Транслокация протеинкиназы С к мембране позволяет ферменту связаться с ДАГ, который ещё больше повышает сродство протеинкиназы С к ионам кальция (рис. 5-38). Наиболее распространённые изоформы протеинкиназы С активируются Са 2+ , диацилглицеролом и фосфатидилсерином.

5-38. Регуляция активности протеинкиназы С (ПКС). ФС - фосфатидилсерин; ДАГ - диаципглицерол.
Каталитический домен имеет центр, связывающий АТФ и белок-субстрат. Активная фермента протеинкиназы С фосфорилирует по остаткам серина и треонина. Снижение концентрации ионов кальция в клетке нарушает связь протеинкиназы С с фосфатидилсерином и диацилглицеролом, фермент переходит в неактивную форму и отделяется от мембраны.
3. Протеинкиназы G
В отличие от протеинкиназы А, протеинкиназа G присутствует не во всех тканях, ее обнаруживают в лёгких, мозжечке, гладких мышцах и тромбоцитах. Изоформы протеинкиназы G могут быть связаны с мембраной или находиться цитоплазме. Растворимая протеинкиназа С состоит из двух идентичных субъединиц, каждая из которых имеет два центра для связывания цГМФ. Приединение цГМФ к регуляторным центрам вызывает конформационные изменения субъединиц и повышает каталитическую активность фермента (рис. 5-39). Протеинкиназа G, подобно протеинкиназе А и С, специфична в отношении определённых белковых субстратов, которые она фосфорилирует по остаткам серина и треонина.

Рис. 5-39. Регуляция активности протеинкиназы G (ПКG).
Ж фосфодиэстеразы
Фосфодиэстеразы - ферменты, катализирующие превращение цАМФ (рис. 5-40) или цГМФ в неактивные метаболиты АМФ или ГМФ. Фосфодиэстеразы, снижая концентрации вторичных посредников, разрывают цепь превращений, вызванных активатором рецептора.

Рис.5-40. Превращение цАМФ в АМФ.
Фосфодиэстеразы присутствуют в клетках тканей в 2 формах: в форме растворимого белка и мембранносвязанного. Формы фермента, связанные с мембраной, в разных тканях составляют 5-40%. В одной и той же ткани могут присутствовать разные формы фосфодиэстеразы, различающиеся по сродству к субстратам, молекулярному весу, заряду, регуляторным свойствам и локализации в клетке.
Фосфодиэстеразы циклических нуклеотидов не обладают абсолютной специфичностью, поэтому, как правило, одна и та же форма фермента способна гидролизовать как цАМФ, так и цГМФ. Однако скорости гидролиза этих двух нуклеотидов под действием одной и той же фосфодиэстеразы могут значительно различаться. Это зависит от того, какая фосфодиэстераза присутствует в клетке - более специфичная в отношении цАМФ или более специфичная к цГМФ, от соотношения концентраций цАМФ и цГМФ в клетке и от действия регуляторов фосфодиэстеразы.
В большинстве тканей присутствует фосфодиэстераза-1, более специфичная к цАМФ, активируемая Са 2+ , комплексом 4 Са 2+ -кальмодулин и цГМФ.
При передаче сигнала на рецепторы, сопряженные , участвует фактически такой же основной механизм. Связывание агониста рецептором приводит к изменению конформации белка рецептора. Это изменение передается на белок G: а-субъединица изменяет гуанозиндифосфат (ГДФ) на гуанозинтрифосфат (ГТФ), затем диссоциирует от двух других субъединиц, связывается с эффекторным белком и изменяет его функциональное состояние.
В принципе, β- и γ-субъединицы тоже способны взаимодействовать с эффекторными белками, а-субъединица медленно гидролизует связанный ГТФ до ГДФ . G a -ГДФ не обладает аффинитетом к эффекторному белку и вновь связывается с β- и γ-субъединицами. Белки G могут диффундировать вдоль мембраны; они не прикреплены к отдельным белкам рецепторов. Тем не менее существует связь между типами рецепторов и типами белка G.
Более того, α-субъединицы отдельных белков G различаются аффинитетом к разным эффекторным белкам, а также характером воздействия на них. G a -ГТФ белка G s стимулирует аденилатциклазу, в то время как G a -ГТФ белка G i является ее ингибитором.
Группа сопряженных с G-белком рецепторов включает мускариновые холинорецепторы, адренорецепторы норадреналина и адреналина, а также рецепторы дофамина, гистамина, серотонина, глутамата, ГАМК, морфина, простагландинов, лейкотриенов и многих других медиаторов и гормонов.
К основным эффекторным белкам у сопряженных с G-белком рецепторов относятся аденилатциклаза (АТФ => внутриклеточный посредник цАМФ), фосфолипаза С (фосфатидилинозитол => внутриклеточные посредники инозитолтрифосфата и диацилглицерола), а также белки ионного канала. Множество клеточных функций регулируется концентрацией клеточного циклического аденозинмонофосфата (цАМФ), поскольку цАМФ увеличивает активность протеинкиназы А, которая катализирует превращение фосфатных групп в функциональные белки.
Повышение уровня цАМФ приводит к снижению тонуса гладких мышц, увеличению сократимости сердечной мышцы, а также повышению гликогенолиза и липолиза. Фосфорилирование белков сердечных Са 2+ -каналов повышает вероятность открытия каналов во время деполяризации мембраны. Следует отметить, что цАМФ инактивируется фосфодиэстеразой. Ингибиторы данного фермента повышают внутриклеточную концентрацию цАМФ и действуют как адреналин.
Белок рецептора самостоятельно фосфорилируется с потерей свойства активировать связанный белок G. Это один из механизмов, который способствует снижению чувствительность клетки во время длительной стимуляции рецептора агонистом (десенситизация).
Активация фосфолипазы С приводит к гидролизу мембранного фосфолипида фосфатидилинозитол-4,5-дифосфата до инозитолтрифосфата (IР 3) и диацил-глицерола (DAG). Инозитолтрифосфат стимулирует высвобождение Са 2+ из органелл-депо, что приводит к сокращению гладкомышечных клеток, распаду гликогена или к активации экзоцитоза. Диацилглицерол стимулирует протеинкиназу С, которая фосфорилирует ряд ферментов, содержащих серин или треонин.
Некоторые белки G вызывают открытие белков каналов. Таким образом активируются К + -каналы (действие АХ на синусовый узел, действие опиоидов на передачу нервного импульса).

Среди органических веществ белки , или протеины , - самые многочисленные, наиболее разнообразные и имеющие первостепенное значение биополимеры. На их долю приходится 50 - 80% сухой массы клетки.
Молекулы белков имеют большие размеры, поэтому их называют макромолекулами . Кроме углерода , кислорода , водорода и азота , в состав белков могут входить сера, фосфор и железо. Белки отличаются друг от друга числом (от ста до нескольких тысяч), составом и последовательностью мономеров. Мономерами белков являются аминокислоты (рис. 1)
Бесконечное разнообразие белков создается за счет различного сочетания всего 20 аминокислот. Каждая аминокислота имеет свое название, особое строение и свойства. Их общую формулу можно представить в следующем виде:
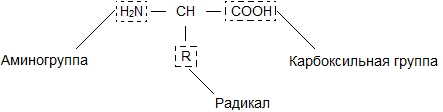
Молекула аминокислоты состоит из двух одинаковых для всех аминокислот частей, одна из которых является аминогруппой (-NH 2 ) с основными свойствами, другая - карбоксильной группой (-COOH ) с кислотными свойствами. Часть молекулы, называемая радикалом (R ), у разных аминокислот имеет различное строение. Наличие в одной молекуле аминокислоты основной и кислотной групп обусловливает их высокую реакционную способность. через эти группы происходит соединение аминокислот при образовании белка. При этом возникает молекула воды, а освободившиеся электроны образуют пептидную связь. Поэтому белки называют полипептидами .
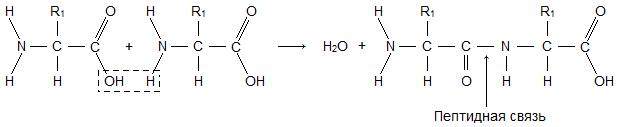
Молекулы белков могут иметь различные пространственные конфигурации, и в их строении различают четыре уровня структурной организации.
Последовательность аминокислот в составе полипептидной цепи представляет первичную структуру
белка. Она уникальна для любого белка и определяет его форму, свойства и функции.
Большинство белков имеют вид спирали в результате образования водородных связей между -CO-
и -NH-
группами разных аминокислотных остатков полипептидной цепи. Водородные связи малопрочные, но в комплексе они обеспечивают довольно прочную структуру. Эта спираль - вторичная структура
белка.
Третичная структура - трехмерная пространственная «упаковка» полипептидной цепи. В результате возникает причудливая, но для каждого белка специфическая конфигурация - глобула . Прочность третичной структуры обеспечивается разнообразными связями, возникающими между радикалами аминокислот.
Четвертичная структура
характерна не для всех белков. Она возникает в результате соединения нескольких макромолекул с третичной структурой в сложный комплекс. Например, гемоглобин крови человека представляет комплекс из четырех макромолекул белка.
Такая сложность структуры белковых молекул связана с разнообразием функций, свойственных этим биополимерам.
Нарушение природной структуры белка называют денатурацией
. Она может происходить под воздействием температуры, химических веществ, лучистой энергии и других факторов. При слабом воздействии распадается только четвертичная структура, при более сильном - третичная, а затем - вторичная, и белок остается в виде полипептидной цепи.
Этот процесс частично обратим: если не нарушена первичная структура, то денатурированный белок способен восстанавливать свою структуру. Отсюда следует, что все особенность строение макромолекулы белка определяются его первичной структурой.
Кроме простых белков , состоящих только из аминокислот, есть еще и сложные белки
G – белки усиливают передаваемый сигнал. Например, передатчик нервного импульса норэпинефрин может взаимодействовать со своим мембранным рецептором всего несколько милисекунд. G – белок увеличивает длительность действия сигнала с милисекунд до десятков секунд, что чрезвычайно важно (не нужно постоянно посылать сигналы нервной системе). Происходит экономия нервной энергии.
Рецепторы, сопряженные с G – белками образуют семейство “серпантинных” (или змеиных) рецепторов, называемых так потому, что их полипептидные цепи пересекают плазматическую мембрану 7 раз.
К этому семейству принадлежат рецепторы для адренергических аминов, серотонина, ацетилхолина (мускариновые), многих пептидных гормонов, обонятельного эпителия, зрительных рецепторов (в колбочках и палочках сетчатки). Информационная молекула (например, норэпинефрин) связывается с «карманом», образованным трансмембранными областями рецептора. Возникшие изменения конформации этих участков передаются цитоплазматическим петлям рецептора, которые активируют G-белок. Чем больше молекул агониста, тем больше скорость его связывания с рецептором.
Десенситизация рецепторов.
Это означает, что после достижения начального высокого уровня эффекта (например, накопление внутриклеточного цАМФ, ток Na + , сокращение мышцы и т.д.) ответ клетки постепенно уменьшается в течение секунд или минут, даже не смотря на постоянное присутствие сигнальной молекулы. Десенситизация обратима. Так, через 15 минут после удаления сигнальной молекулы, его повторное воздействие ведет к реакции, сравнимой по величине с начальной.
Down – регуляция рецепторов.
Рецептор, при его чрезмерной стимуляции, может погружаться в цитозоль и клетка с помощью лизосомальных ферментов «переваривает» его до аминокислот. Мембрана, где был рецептор, восстанавливается.
Ар – регуляция.
Если хирургически перерезать нервы, иннервирующие мышцу, то мышца не получит сигнал из нервной системы и не может сократиться. Реакция мышцы на денервацию направлена на синтез дополнительных рецепторов. Они синтезируются и встраиваются в наружную клеточную мембрану. Клетка хочет получить сигнал на сокращение. Сигнал не поступает (нерв перерезан), хотя рецепторов много и они особенно восприимчивы к нейромедиатору. Рецепторы располагаются даже в других местах, вдали от места соединения нерва с мышцей. Это и есть так называемая Ар – регуляция рецепторов – синтез новых рецепторов клеткой и встраивание их в мембрану. Рецепторы постоянно обновляются. Срок жизни рецептора – несколько дней. Взамен состарившемуся и разрушенному клеткой, она строит новый. Это динамичный процесс.
Таким образом, встроенный в мембрану рецептор принимает сигнал (нервный импульс, гормон лекарства), G-белок усиливает этот сигнал. Эффекторный элемент (фермент) реализует этот сигнал, запуская синтез в клетке вторичных посредников. Они изменяют скорость протекания биохимических реакций в клетках и непосредственно реализуют сигнал, посылаемый нервной или гормональной системой.
Вторичные посредники.
1) цАМФ. участвует в передаче таких гормональных эффектов, как: 1) мобилизация энергетических запасов (распад углеводов в печени или триглицеридов в жировых клетках – эффекты катехоламинов – (эпинефрин, изопреналин).
2) задержка воды почками – эффекты вазопресина;
3) поддержание Са +2 гомеостаза – эффекты гормонов паращитовидных желез;
4) увеличение частоты и силы сокращений сердечной мышцы – эффекты катехоламинов (эпинефрин, изопреналин)
5) регуляция биосинтеза стероидов в надпочечниках и половых железах – эффекты кортикотропина или фолликулостимулирующего гомона;
6) – расслабление гладких мышц и многие другие гормональные и нервные процессы.
Когда нервный или гормональный стимул завершается, внутриклеточные эффекты цАМФ прекращаются путем активации фермента разрушающего цАМФ.
Одним из механизмов лечебного действия кофеина , теофиллина и других метилксантинов является ингибирование распада цАМФ.
2) Са +2 и фосфоинозитиды.
Некоторые гормоны, нейромедиаторы и факторы роста связываются с рецептором на поверхности эффекторной клетки. Сигнал передается на G-белок. В последующем происходит активация фосфолипазы С. Последняя специфически расщепляет фосфолипиды плазматической мембраны с образованием двух вторичных посредников: 1) диацилглицерола , 2) инозитолтрифосфата .
Диацилглицерол активирует протеинкиназу С, которая фосфорилирует ферменты и изменяет их активность.
Инозитолтрифосфат высвобождает Са 2+ из внутриклеточных хранилищ (саркоплазматический ретикулум, митохондрии). Са 2+ изменяет функции клетки. Например провоцирует сокращение мышцы и т.д.).
Через фосфоинозитиды действует литий, используемый для лечения маниакально-депрессивных состояний.
3) цГМФ . В отличие от цАМФ, участвует в передаче сигналов лишь в некоторых типах клеток. В слизистой кишечника и гладких мышцах сосудов функционирует параллельно с цАМФ-системой (как запасной). Механизм действия цГМФ также опосредован фосфорилированием белков.
Повышенная концентрация цГМФ вызывает расслабление гладкой мускулатуры сосудов за счет дефосфорилирования легких цепей миозина.
Фосфорилирование: общий механизм.
Почти все механизмы передачи сигнала с помощью вторичных посредников обусловлены фосфорилированием.
В процессе эволюции организм не выработал специальные рецепторы для лекарств. Они действуют через рецепторы для нейромедиаторов и гормонов. Почти все лекарства (исключение составляют, пожалуй, лишь средства для общей анестезии) оказывают свое действие через рецепторы.
Мы детально рассмотрели рецепторы, встроенные в плазматическую мембрану клетки. Но есть и другие рецепторы лекарств. В принципе рецептор – это то, с чем лекарство связывается (взаимодействует) в организме. Например, альбумин – рецептор для лекарств, которые связываются с ним. Но этот рецептор не активный, не приводящий к возникновению фармакологического эффекта.
К другим классам рецепторов лекарств относятся:
1) ферменты, 2) транспортные белки, 3) структурные белки.
При связывании с лекарствами они могут ингибироваться или (реже) активироваться. Например, дигидрофолатредуктаза – рецептор для метотрексата.
Транспортные белки (например, мембранный рецептор для сердечных гликозидов – Nа + , К + , АТФаза).
Структурные белки (например, тубулин – рецептор для противовоспалительного средства колхицина) .
В каждом случае взаимодействия лекарства с рецептором образуется лекарственно-рецепторный комплекс, приводящий к изменению метаболизма в клетке и органе. Развивается фармакологический эффект. Его величина пропорциональна количеству лекарственно-рецепторных комплексов.
Лекарства, действие которых связано с возбуждением рецепторов называют агонистами . Агонисты бывают: 1) полные (вызывают максимальный ответ) и 2) частичные. Последние связываются с рецепторами и возбуждают их. Но фармакологический эффект слабее, чем от природного регулятора. Вещества, препятствующие действию специфических агонистов называются антагонистами (блокаторами).
Рецепторы классифицируют по их чувствительности к естественным медиаторам и к их антагонистам. Например, чувствительные к ацетилхолину рецепторы называют холинэргическими, чувствительные к эпинефрину (адреналину) – адренергическими.
Для возбуждения рецепторов и получения соответствующего эффекта используют как сами медиаторы (норэпинефрин, дофамин и другие), так и лекарства, обладающие сродством к рецепторам. Чаще всего последние являются структурными аналогами медиаторов.
Некоторые вещества возбуждают соответствующий рецептор не путем непосредственного взаимодействия с ним, а за счет освобождения медиаторов из связанной (физиологически неактивной) формы или путем угнетения ферментов, разрушающих медиаторы.
Рецепторы занимают небольшую часть наружной клеточной мембраны. Так, участки мембраны, реагирующие на ацетилхолин, составляют только 1/6000 часть общей поверхности клетки
2. ВЛИЯНИЕ НА АКТИВНОСТЬ ФЕРМЕНТОВ. Действие некоторых лекарств основано на активации или ингибировании ферментов. Например, ФИЗОСТИГМИН угнетает активность холинэстеразы, разрушающей ацетилхолин. Он вызывает эффекты, характерные для возбуждения парасимпатической нервной системы.
Некоторые лекарства способны вызывать индукцию, то есть увеличивать содержание ферментного белка. При этом возрастает их активность. Например, фенобарбитал, повышая активность УДФ-глюкуронилтрансферазы снижают гипербилирубинемию.
3. ФИЗИКО-ХИМИЧЕСКОЕ ВОЗДЕЙСТВИЕ НА МЕМБРАНЫ КЛЕТОК .
Для некоторых лекарств природа молекул-мишеней неизвестна. Их действие не связано со специфическими рецепторами. Например, средства для общей анестезии действуют, изменяя транспорт ионов. Лечебный эффект мазей, присыпок, жидких мазей имеет физическую природу. Они предохраняют пораженные участки кожи или слизистых оболочек от раздражения.
Лекарства, действующие на рецепторы , вызывают широкий спектр тканевых и системных ответов по двум причинам. Во-первых, на разных тканях находятся различные рецепторы. Во-вторых, различным типам рецепторов присуща разная структура и, соответственно, функция, поэтому клеточный ответ на активацию рецептора (трансдукция) значительно варьирует в зависимости от структуры рецептора. Выделяют четыре типа рецепторов, которые отличаются кинетическими характеристиками:
G-белок-связанные рецепторы;
ДНК-связанные рецепторы;
рецепторы, обладающие тирозинкиназной активностью (тирозинкиназные рецепторы);
Рецептор-связанные каналы (РСК).
Тирозинкиназные рецепторы и РСК отличаются от остальных тем, что для проявления клеточного ответа они не требуют клеточной трансдукции компонентов при активации агонистом, поэтому молекула рецептора играет большую роль, чем молекулярное звено между лекарством и трансдукцией. Тирозинкиназный рецептор - в действительности мембраносвязанный фермент, который «включается» агонистом. РСК - специфический ионный канал, который структурно отличается от потенциалзависимых каналов, регулируемых лекарством или нейротрансмиттером, тем, что он присоединяется к высокоспецифическому лиганд-связывающему центру, а не к специфическому потенциалчувствительному участку. В РСК лиганд-связывающий центр и канал являются функционально различными участками одной молекулы. РСК также называют лиганд-зависимым ионным каналом. Приведенная классификация удобна, поскольку рассматривает тирозинкиназные структуры и РСК как рецепторы.
G-белок-связанные рецепторы
G-белки являются компонентами трансдукции. Рецепторы, связанные с G-белками, локализованы на клеточной мембране и состоят из 7 трансмембранных спиралей (I-VII).
В отсутствие агониста рецептор связан с G-белком, что удерживает рецептор в неактивной конформации. Сам по себе - это комплекс трех субъединиц (а, бета и у). Когда рецептор находится в состоянии покоя, три субъединицы G-белка и гуанозиндифосфат (ГДФ) прочно связаны с альфа-субъединицей G-белка. Связывание агониста приводит к конформационным изменениям рецептора, что, в свою очередь, вызывает конформационное изменение G-белка, ведущее к диссоциации ГДФ от альфа-субъединицы. Это инициирует последовательность реакций и способствует трансдукции G-белок-связанного рецептора, которая описывается далее.
Из компонентов трансдукции , непосредственно связанных с G-белками, наиболее распространенным в организме является аденилилциклаза. Циклический нуклеотид цАМФ синтезируется из аденозинтрифосфата (АТФ) ферментом аденилилциклазой. цАМФ обладает различными биологическими эффектами.
цАМФ оказывает действие на энергетический обмен, клеточную дифференцировку, функцию ионных каналов и сократительные белки
цАМФ фосфорилирует внутриклеточные белки (чаще ферменты) через воздействие цАМФ-зависимых протеинкиназ. Эти протеинкиназы активируют цАМФ и фосфорилируют аминокислоты серии и треонин, используя АТФ как источник фосфора. Результат фосфорилирования состоит в:
Активации гормон-чувствительных липаз;
инактивации гликогенсинтазы;
активации фосфорилазкиназы и превращении неактивной фосфорилазы в активную, что приводит к ускорению липолиза, снижению синтеза и ускорению распада гликогена;
активации Са2+-каналов L-типа и саркоплазматического ретикулума в сердечных клетках за счет фосфорилирования, что увеличивает выход кальция.










