Что такое нуклеотид в биологии. Из чего состоит нуклеотид и что это такое. Уровни организации генома эукариот
Нуклеотиды.
Нуклеиновые кислоты
Нуклеотиды
Нуклеотиды – это природные соединения, состоящие из 1) остатков азотистого нуклеинового основания, 2) углеводного остатка и 3) фосфатной группы.
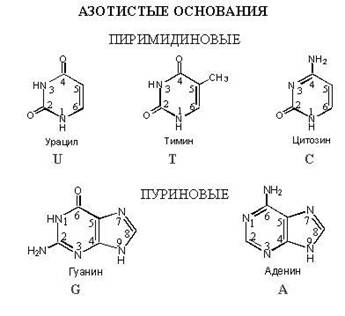
Азотистые нуклеиновые основания
Азотистые основания – это производные двух гетероциклов - пиримидина и пурина .
Пиримидиновые основания

Пуриновые основания

ТАУТОМЕРИЯ АЗОТИСТЫХ ОСНОВАНИЙ
а) лактам-лактимная

Аналогичная таутомерия возможна у тимина, цитозина и гуанина.
б) амино-иминная

Аналогичная таутомерия возможна у гуанина и цитозина.
Лактамы более устойчивы, чем лактимы, а амины более устойчивы, чем имины. Все основания in vitro и in vivo существуют и участвуют в обмене веществ в лактамных и аминоформах.
Производные и аналоги нуклеиновых оснований применяются в медицине как лекарственные вещества противоопухолевого действия:

Нуклеозиды
Нуклеозиды – это соединения, состоящие из остатков нуклеинового основания и углевода, связанных β- N -гликозидной связью.

Реакция образования нуклеозидов in vivo идет под действием ферментов.
В кислой среде (но не в нейтральной и не в щелочной) нуклеозиды гидролизуются, распадаясь на исходные основание и углевод. Пиримидиновые нуклеозиды гидролизуются труднее, пуриновые – легче.


Номенклатура нуклеозидов
|
Основание |
Название |
|
|
2"-Дезоксиуридин |
||
|
2"-Дезокситимидин |
||
|
2"-Дезоксицитиидин |
||
|
Аденозин |
||
|
2"-Дезоксиаденозин |
||
|
Гуанозин |
||
|
2"-Дезоксигуанозин |
Нуклеотиды
Нуклеотиды – это нуклеозиды, содержащие фосфатную группу в 5"-положении (5"-фосфоридированные нуклеозиды).
Нуклеотиды образуются in vivo в результате ферментативного фосфорилирования нуклеозидов:

Нуклеотиды гидролизуются в кислой и в щелочной средах: при кислотном гидролизе образуются основание, углевод и фосфорная кислота, а щелочной гидролиз дает нуклеозид и фосфат натрия:

Номенклатура нуклеотидов
|
Основание |
Название |
|
|
Уридин-5"-монофосфат (УМФ), уридиловая кислота |
||
|
2"-Дезоксиуридин-5"-монофосфат |
||
|
Тимидин-5"-монофосфат (ТМФ), тимидиловая кислота |
||
|
2"-Дезокситимидин-5"-монофосфат |
||
|
Цитидин-5"-монофосфат (ЦМФ), цитидиловая кислота |
||
|
2"-Дезоксицитиидин-5"-монофосфат |
||
|
Аденозин-5"-монофосфат (АМФ), адениловая кислота |
||
|
2"-Дезоксиаденозин-5"-монофосфат |
||
|
Гаунозин-5"-монофосфат (ГМФ), гуаниловая кислота |
||
|
2"-Дезоксигуанозин-5"-монофосфат |
Динуклеотиды


НАД и ФАД – коферменты, участвующие в ОВ реакциях переноса водорода в организме:


Аденозинтрифосфат (АТФ)

АТФ является аккумулятором и переносчиком энергии в биохимических реакциях.
Биологические реакции АТФ
1. Фосфорилирование – перенос фосфатных групп от АТФ на другие субстраты:

2. Гидролиз с выделением энергии, используемой в синтетазных реакциях:

Нуклеиновые кислоты
Нуклеиновые кислоты – это полинуклеотиды – полимеры, состоящие из нуклеотидных остатков, связанных сахарофосфатными сложноэфирными связями.
Схема строения полинуклеотидной цепи:

Виды НК : ДНК – содержат остатки 2 -дезоксирибозы, не содержат урацила;
Д Н К
Первичная структура ДНК
Первичная структура ДНК – это определенный порядок последовательности нуклеотидов в цепи:

В первичной структуре ядерной ДНК заложен генетический код. В процессе транскрипции он "переписывается" на информационную РНК, а затем происходит трансляция: в рибосоме на матрице информационной РНК синтезируется полипептидная цепь белка. Ключ генетического кода заключается в том, что один аминокислотный остаток в синтезируемой полипептидной цепи кодируется тремя нуклеотидными остатками (триплетом) в НК, и таким образом, с помощью 4 видов нуклеотидов кодируются 20 аминокислот.
Химические свойства нуклеиновых кислот
Сложноэфирные связи, соединяющие полинуклеотидные цепи, неустойчивы в кислой и щелочной средах, и НК в этих условиях подвергаются гидролизу:

КОМПЛЕМЕНТАРНОСТЬ АЗОТИСТЫХ ОСНОВАНИЙ
Комплементарность – это соответствие формы двух сложных линий, которые подходят друг к другу "как ключ к замку".
Комплементарные пары оснований:

В паре А-Т тимин может быть (при переходе ДНК→РНК) заменен урацилом, и пара становится А-У ("взаимозаменяемость" тимина и урацила).
Биологическое значение комплементарных взаимодействий заключается в том, что они обеспечивают точность передачи информации от одной НК к другой.
Вторичная структура ДНК
Представляет собой спираль, состоящую из двух комплементарных друг другу и антипараллельных полинуклеотидных цепей ("двойная спираль"):

Биологическая роль "двойной спирали":
1) Она обеспечивает сохранность генетической информации (ядерный нуклеопротеидный комплекс "ДНК-Гистоны");
2) Обеспечивает восстановление информации при повреждении ДНК (репарация после мутаций).
Р Н К
Виды РНК : рибосомальная, информационная, транспортная.
Рибосомальная РНК (р-РНК) – структурный материал рибосом (рибосомальный нуклеопротеидный клмплекс).
Информационная (матричная) РНК (и-РНК) – промежуточный этап в процессе трансформации информации "ДНК – белок". Она синтезируется на матрице ДНК и сама служит матрицей при синтезе белка в рибосоме. и-РНК сравнительно низкомолекулярна и не имеет развитой вторичной структуры.
Транспортная РНК (т-РНК) – низкомолекулярная РНК, выполняющая следующие функции: 1) определение "своей" аминокислоты (для каждой АК существует своя т-РНК); 2) связывание с АК и транспорт её к рибосоме; 3) определение места АК в растущей полипептидной цепи.
Транспортные РНК имеют вторичную структуру "клеверного листа":

Выступающий конец ССА-3 ОН – место связывания с карбоксильной группой АК.
Триплет нуклеотидов в крайней нижней точке – кодон, комплементарный соответствующему антикодону на и-РНК.
ЛИТЕРАТУРА:
Основная
1. Тюкавкина Н.А., Зурабян С.Э., Белобородов В.Л. и др. – Органическая химия (специальный курс), кн.2 – Дрофа, М., 2008 г., с. 157-178.
2. Н.А.Тюкавкина, Ю.И.Бауков – Биоорганическая химия – ДРОФА, М., 2007 г., с. 420-444.
Все живое на планете состоит из множества клеток, поддерживающих упорядоченность своей организации за счет содержащейся в ядре генетической информации. Она сохраняется, реализуется и передается сложными высокомолекулярными соединениями - нуклеиновыми кислотами, состоящими из мономерных звеньев - нуклеотидов. Роль нуклеиновых кислот невозможно переоценить. Стабильностью их структуры определяется нормальная жизнедеятельность организма, а любые отклонения в строении неминуемо приводят к изменению клеточной организации, активности физиологических процессов и жизнеспособности клеток в целом.
Понятие нуклеотида и его свойства
Каждая или РНК собрана из более мелких мономерных соединений - нуклеотидов. Другими словами, нуклеотид - это строительный материал для нуклеиновых кислот, коферментов и многих других биологических соединений, которые крайне необходимы клетке в процессе ее жизнедеятельности.
К основным свойствам этих незаменимых веществ можно отнести:
Хранение информации о и наследуемых признаках;
. осуществление контроля над ростом и репродукцией;
. участие в метаболизме и многих других физиологических процессах, протекающих в клетке.
Говоря о нуклеотидах, нельзя не остановиться на таком важном вопросе, как их структура и состав.

Каждый нуклеотид состоит из:
Сахарного остатка;
. азотистого основания;
. фосфатной группы или остатка фосфорной кислоты.
Можно сказать, что нуклеотид - это сложное органическое соединение. В зависимости от видового состава азотистых оснований и типа пентозы в структуре нуклеотида нуклеиновые кислоты подразделяются на:
Дезоксирибонуклеиновую кислоту, или ДНК;
. рибонуклеиновую кислоту, или РНК.
Состав нуклеиновых кислот
В нуклеиновых кислотах сахар представлен пентозой. Это пятиуглеродный сахар, в ДНК его называют дезоксирибозой, в РНК - рибозой. Каждая молекула пентозы имеет пять атомов углерода, четыре из них вместе с атомом кислорода образуют пятичленное кольцо, а пятый входит в группу НО-СН2.
Положение каждого атома углерода в молекуле пентозы обозначается арабской цифрой со штрихом (1C´, 2C´, 3C´, 4C´, 5C´). Поскольку все процессы считывания с молекулы нуклеиновой кислоты имеют строгую направленность, нумерация атомов углерода и их расположение в кольце служат своего рода указателем правильного направления.
По гидроксильной группе к третьему и пятому углеродным атомам (3С´ и 5С´) присоединен остаток фосфорной кислоты. Он и определяет химическую принадлежность ДНК и РНК к группе кислот.
К первому углеродному атому (1С´) в молекуле сахара присоединено азотистое основание.
Видовой состав азотистых оснований
Нуклеотиды ДНК по азотистому основанию представлены четырьмя видами:
Аденином (А);
. гуанином (Г);
. цитозином (Ц);
. тимином (Т).
Первые два относятся к классу пуринов, два последних - пиримидинов. По молекулярной массе пуриновые всегда тяжелее пиримидиновых.

Нуклеотиды РНК по азотистому основанию представлены:
Аденином (А);
. гуанином (Г);
. цитозином (Ц);
. урацилом (У).
Урацил так же, как и тимин, является пиримидиновым основанием.
В научной литературе нередко можно встретить и другое обозначение азотистых оснований - латинскими буквами (A, T, C, G, U).
Подробнее остановимся на химической структуре пуринов и пиримидинов.

Пиримидины, а именно цитозин, тимин и урацил, в своем составе представлены двумя атомами азота и четырьмя атомами углерода, образующих шестичленное кольцо. Каждый атом имеет свой номер от 1 до 6.
Пурины (аденин и гуанин) состоят из пиримидина и имидазола или двух гетероциклов. Молекула пуриновых оснований представлена четырьмя атомами азота и пятью атомами углерода. Каждый атом пронумерован от 1 до 9.
В результате соединения азотистого основания и остатка пентозы образуется нуклеозид. Нуклеотид - это соединение нуклеозида и фосфатной группы.
Образование фосфодиэфирных связей
Важно разобраться в вопросе о том, как соединяются нуклеотиды в полипептидную цепь и образуют молекулу нуклеиновой кислоты. Происходит это за счет так называемых фосфодиэфирных связей.
Взаимодействие двух нуклеотидов дает динуклеотид. Образование нового соединения происходит путем конденсации, когда между фосфатным остатком одного мономера и гидроксигруппой пентозы другого возникает фосфодиэфирная связь.
Синтез полинуклеотида - неоднократное повторение этой реакции (несколько миллионов раз). Полинуклеотидная цепь строится посредством образования фосфодиэфирных связей между третьим и пятым углеродами сахаров (3С´ и 5С´).
Сборка полинуклеотида - сложный процесс, протекающий при участии фермента ДНК-полимеразы, которая обеспечивает рост цепи только с одного конца (3´) со свободной гидроксигруппой.
Структура молекулы ДНК
Молекула ДНК, так же как и белка, может иметь первичную, вторичную и третичную структуру.

Последовательность нуклеотидов в цепи ДНК определяет ее первичную формируется за счет водородных связей, в основе возникновения которых положен принцип комплементарности. Другими словами, при синтезе двойной действует определенная закономерность: аденин одной цепи соответствует тимину другой, гуанин - цитозину, и наоборот. Пары аденина и тимина или гуанина и цитозина образуются за счет двух в первом и трех в последнем случае водородных связей. Такое соединение нуклеотидов обеспечивает прочную связь цепей и равное расстояние между ними.
Зная последовательность нуклеотидов одной цепи ДНК, по принципу комплементарности или дополнения можно достроить вторую.
Третичная структура ДНК образована за счет сложных трехмерных связей, что делает ее молекулу более компактной и способной размещаться в малом объеме клетки. Так, например, длина ДНК кишечной палочки составляет более 1 мм, тогда как длина клетки - меньше 5 мкм.
Число нуклеотидов в ДНК, а именно их количественное соотношение, подчиняется правилу Чергаффа (число пуриновых оснований всегда равно количеству пиримидиновых). Расстояние между нуклеотидами - величина постоянная, равная 0,34 нм, как и их молекулярная масса.
Структура молекулы РНК
РНК представлена одной полинуклеотидной цепочкой, образованной через между пентозой (в данном случае рибозой) и фосфатным остатком. По длине она значительно короче ДНК. По видовому составу азотистых оснований в нуклеотиде также имеются различия. В РНК вместо пиримидинового основания тимина используется урацил. В зависимости от функций, выполняемых в организме, РНК может быть трех типов.

Рибосомальная (рРНК) - содержит обычно от 3000 до 5000 нуклеотидов. Как необходимый структурный компонент принимает участие в формировании активного центра рибосом, места осуществления одного из важнейших процессов в клетке — биосинтеза белка.
. Транспортная (тРНК) - состоит в среднем из 75 - 95 нуклеотидов, осуществляет перенос нужной аминокислоты к месту синтеза полипептида в рибосоме. Каждый вид тРНК (не менее 40) имеет свою, присущую только ему последовательность мономеров или нуклеотидов.
. Информационная (иРНК) - по нуклеотидному составу весьма разнообразна. Переносит генетическую информацию от ДНК к рибосомам, выступает в роли матрицы для синтеза белковой молекулы.
Роль нуклеотидов в организме
Нуклеотиды в клетке выполняют ряд важнейших функций:
Используются в качестве структурных блоков для нуклеиновых кислот (нуклеотиды пуринового и пиримидинового рядов);
. участвуют во многих обменных процессах в клетке;
. входят в состав АТФ - главного источника энергии в клетках;
. выступают в роли переносчиков восстановительных эквивалентов в клетках (НАД+, НАДФ+, ФАД, ФМН);
. выполняют функцию биорегуляторов;
. могут рассматриваться как вторые вестники внеклеточного регулярного синтеза (например, цАМФ или цГМФ).
Нуклеотид - это мономерная единица, образующая более сложные соединения - нуклеиновые кислоты, без которых невозможна передача генетической информации, ее хранение и воспроизведение. Свободные нуклеотиды являются главными компонентами, участвующими в сигнальных и энергетических процессах, поддерживающих нормальную жизнедеятельность клеток и организма в целом.
Остановимся подробнее на нуклеотидах. Известно, что нуклеотиды называются аденин, гуанин, тимин, цитозин и урацил – азотистые основания, они представлены на рисунке ниже.
Нуклеотиды – это мономеры нуклеиновых кислот. Нуклеиновые кислоты в эукариотических клетках находятся в ядре. Они есть у всех живых организмов (у тех, у кого нет ядра, нуклеиновые кислоты все равно есть – они находятся в центре клетки у бактерий и образуют нуклеоиды). Мономеры, из которых потом строятся нуклеиновые кислоты, состоят из азотистого основания, остатка сахара (дезоксирибоза или рибоза) и фосфата. Сахара вместе с азотистым основанием называются нуклеозидами (аденозин, гуанозин, тимидин, цитидин). Если к ним присоединены 1-, 2-, или 3-фосфорных остатка, то вся эта структура называется Соответственно, нуклеотизид монофосфатом, дифосфатом или трифосфатом или нуклеотидом (аденин, гуанин, тимин, цитозин).

Вот так модель АТФ выглядит в пространстве. Азотистое основание, входящее в состав ДНК делится на две группы – пиримидиновую и пуриновую. В состав ДНК входит аденин, тимин, цитозин и гуанин, в РНК вместо тимина урацил. Как известно, ДНК – это большой архив, в котором хранится информация, а РНК – это молекула, которая переносит информацию из ядра в цитоплазму для синтеза белков. С различием в функциях связаны различия в строении. РНК более химически активно из-за того, что ее сахар - рибоза – имеет в своем составе гидроксильную группу, а в дезоксирибозе кислорода нет. Из-за отсутствия кислорода ДНК более инертно, что важно для ее функции хранения информации, чтобы она не вступала ни в какие реакции.

Нуклеотиды способны взаимодействовать друг с другом, при этом «выбрасывается» два фосфора, и между соседними нуклеотидами образуется связь. В молекуле фуранозы молекулы углерода пронумерованы. С первым связано азотистое основание. Когда образуется цепочка нуклеотидов, связь осуществляется между пятым углеродом одной и третьим углеродом другой фосфорной кислоты. Поэтому в цепочке нуклеиновых кислот выделяют разные неравнозначные концы, относительно которых молекула не симметрична.

Комплементарные друг другу одноцепочечные молекулы нуклеиновой кислоты способны образовывать двуцепочечную структуру. Внутри этой спирали аденин образует пару с тимином, а гуанин - с цитозином. Встречается утверждение, что нуклеотиды подходят друг другу как осколки разбитого стекла, поэтому они и образуют пары. Но это утверждение неверно. Нуклеотиды способны образовывать пары как угодно. Единственная причина, по которой они соединяются так, и никак иначе, заключается в том, что угол между «хвостиками», которые идут к сахарам, совпадает только в этих парах, и, кроме того, совпадают их размеры. Никакая другая пара не образует такой конфигурации. А поскольку они совпадают, то их через сахаро-фосфатный остов можно связать друг с другом. Структуру двойной спирали открыли в 1953 году Джеймс Уотсон и Фрэнсис Крик.

При соединение друг с другом против 5’-конца одной нити находится 3’-конец другой нити. То есть нити идут в противоположных направлениях – говорят, что нити в ДНК антипараллельны.
На рисунке видна модель ДНК, видно, что аденин соединяется с тимином двумя водородными связями, а гуанин соединяется с цитозином тройной водородной связью. Если молекулу ДНК подогревать, то ясно, что две связи легче разорвать, чем три, это существенно для свойств ДНК.

В силу пространственного расположения сахаро-фосфатного остова и нуклеотидов, когда нуклеотиды накладывают один на другой и «сшивают» через сахаро-фосфатный остов, цепочка начинает заворачиваться, тем самым образуя знаменитую двойную спираль.
На рисунках представлены шариковые модели ДНК, где каждый атом обозначен шариком. Внутри спирали имеются бороздки: маленькая и большая. Через эти бороздки с ДНК взаимодействуют белки и распознают там последовательность нуклеотидов.
При нагревании ДНК водородные связи разрываются и нити в двойной спирали расплетаются. Процесс нагревания называется плавлением ДНК, при этом разрушаются связи между парами А-Т и Г-Ц.Чем больше в ДНК пар А-Т, тем менее прочно нити друг с другом связаны, тем легче ДНК расплавить. Переход из двухспиральной ДНК в одно-спиральную измеряется на спектрофотометрах по поглощению света при 260 нм. Температура плавления ДНК зависит от А-Т/Г-Ц состава и размера фрагмента молекулы. Ясно, что если фрагмент состоит из нескольких десятков нуклеотидов, то его гораздо легче расплавить, чем более длинные фрагменты.

У человека в гаплоидном геноме, то есть единичном наборе хромосом, 3 млрд. пар нуклеотидов, и их длина составляет 1,7 м, а клетка гораздо меньше, как вы догадываетесь. Для того, чтобы ДНК смогла в ней поместиться, она достаточно плотно свернута, и в эукариотической клетке свернуться ей помогают белки – гистоны. Гистоны имеют положительный заряд, а так как ДНК заряжена отрицательно, то гистоны обладают сродством к ДНК. Упакованная при помощи гистонов ДНК имеет вид бусин, называемых нуклеосомами. 200 пар нуклеотидов идет на одну нуклеосому, 146 пар накручиваются на гистоны, а остальные 54 висят в виде линкерных (связывающих нуклеосомы) ДНК. Это первый уровень компактизации ДНК. В хромосомах ДНК свернута еще несколько раз для того, чтобы образовались компактные структуры.

К нуклеиновым кислотам кроме ДНК относится также РНК. В клетке присутствуют разные типы РНК: рибосомные, матричные, транспортные. Существуют и другие виды РНК, о которых мы будем говорить позже. РНК синтезируется в виде одно-цепочечной молекулы, но отдельные ее участки входят в состав двуцепочечных спиралей. Для РНК также говорят о первичной структуре (последовательности нуклеотидов) и вторичной структуре (образование двуспиральных участков).
К 1944 г. О. Эйвери и его коллеги К. Маклеод и М. Маккарти открыли трансформирующую активность ДНК у пневмококков. Эти авторы продолжили работу Гриффита, описавшего феномен трансформации (передачи наследственных признаков) у бактерий. О. Эйвери, К. Маклеод, М. Маккарти показали, что при удалении белков, полисахаридов и РНК трансформация бактерий не нарушается, а при воздействии на индуцирующее вещество ферментом дезоксирибонуклеазой трансформирующая активность исчезает.
В этих экспериментах впервые была продемонстрирована генетическая роль молекулы ДНК. В 1952 г. А. Херши и М. Чейз подтвердили генетическую роль молекулы ДН К в опытах на бактериофаге Т2. Пометив его белок радиоактивной серой, а ДНК-радиоактивным фосфором,они инфицировали этим бактериальным вирусом кишечную палочку Е. coli. В потомстве фага было выявлено большое количество радиоактивного фосфора и лишь следы S. Отсюда следовало, что именно ДНК, а не белок фага проникает в бактерию, а затем после репликации передается фаговому потомству.
Строение нуклеотида ДНК. Типы нуклеотидов.
Нуклеотид ДНК состоит из
Азотистого основания (в ДНК 4 типа: аденин, тимин, цитозин, гуанин)
Моносахара дезоксирибозы
Фосфорной кислоты
Молекула нуклеотида состоит из трех частей - пятиуглеродного сахара, азотистого основания и фосфорной кислоты.
Сахар, входящий в состав нуклеотида , содержит пять углеродных атомов, т. е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два типа нуклеиновых кислот - рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу. В дезоксирибозе - ОН-группа при 2-м атоме углерода заменена на атом Н, т. е. в ней на один атом кислорода меньше, чем в рибозе.
В обоих типах нуклеиновых кислот содержатся основания четырех разных видов: два из них относятся к классу пуринов и два - к классу пиримидинов. Основной характер этим соединениям придает включенный в кольцо азот. К числу пуринов относятся аденин (А) и гуанин (Г), а к числу пиримидинов - цитозин (Ц) и тимин (Т) или урацил (У) (соответственно в ДНК или РНК). Тимин химически очень близок к урацилу (он представляет собой 5-метилурацил, т. е. урацил, в котором у 5-го углеродного атома стоит метильная группа). В молекуле пуринов имеется два кольца, а в молекуле пиримидинов - одно.
Нуклеотиды соединяются между собой прочной ковалентной связью через сахар одного нуклеотида и фосфорную кислоту другого. Получаетсяполинуклеотидная цепь . На одном ее конце – свободная фосфорная кислота (5’-конец), на другом – свободный сахар (3’-конец). (ДНК-полимераза может присоединять новые нуклеотиды только к 3’-концу.)
Две полинуклеотидные цепи соединяются друг с другом слабыми водородными связями между азотистыми основаниями. Соблюдаются 2 правила:
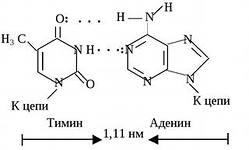
принцип комплементарности: напротив аденина всегда стоит тимин, напротив цитозина – гуанин (они подходят друг другу по форме и числу водородных связей – между А и Г две связи, между Ц и Г – 3).
принцип антипараллельности: там, где у одной полинуклеотидной цепи 5’-конец, у другой – 3’-конец, и наоборот.
Получается двойная цепь ДНК.
Она скручивается в двойную спираль , один виток спирали имеет длину 3,4 нм, содержит 10 пар нуклеотидов. Азотистые основания (хранители генетической информации) находятся внутри спирали, защищенные.
Структурная организация молекулы ДНК. Модель Дж.Уотсона и Ф.Крика
В 1950 г. английский физик М.Уилкинс получил рентгенограмму кристаллических волокон ДНК. Она показала, что молекула ДНК имеет определенную структуру, расшифровка которой помогла бы понять механизм функционирования ДНК. Рентгенограммы, полученные не на кристаллических волокнах ДНК, а на менее упорядоченных агрегатах, которые образуются при более высокой влажности, позволили Розалинд Франклин, коллеге М. Уилкинса, увидеть четкий крестообразный рисунок - опознавательный знак двойной спирали. Стало известно также, что нуклеотиды расположены друг от друга на расстоянии 0,34 нм, а на один виток спирали их приходится 10. Диаметр молекулы ДНК составляет около 2 нм. Из рентгеноструктурных данных, однако, было не ясно, каким образом цепи удерживаются вместе в молекулах ДНК.
Картина полностью прояснилась в 1953 г., когда американский биохимик Дж. Уотсон и английский физик Ф. Крик, исследуя структуру молекулы ДНК, пришли к выводу, что сахарофосфатный остов находится на периферии молекулы ДНК, а пуриновые и пиримидиновые основания - в середине. Причем последние ориентированы таким образом, что между основаниями из противоположных Цепей могут образоваться водородные связи. Из построенной ими модели выявилось, что какой-либо пурин в одной цепи всегда связан водородными связями с одним из пиримидинов в другой цепи. Такие пары имеют одинаковый размер по всей длине молекулы. Не менее важно то, что аденин может спариваться лишь с тимином, а гуанин только с с цитозином. При этом между аденином и тимином образуются две водородные связи, а между гуанином и цитозином – три.
Свойства и функции ДНК.
Хранение наследственной информации (генетический код – способ записи ген.информации о последовательности аминокислот в белке с помощью нуклеотидов (Гамов)
Передача (репликация/удвоение)
Реализация (транскрипция)
Ауторепродукция ДНК. Репликон и его функционирование.
Процесс самовоспроизведения молекул нуклеиновых кислот, сопровождающийся передачей по наследству (от клетки к клетке) точных копий генетической информации; осуществляется с участием набора специфических ферментов (геликаза, контролирующая расплетание молекулы ДНК, ДНК-полимеразы, ДНК-лигаза), проходит по полуконсервативному типу с образованием репликативной вилки; на одной из цепей синтез комплементарной цепи непрерывен, а на другой происходит за счет образования фрагментов Дказаки. Высокоточный процесс, частота ошибок при котором не превышает 10 -9 ; у эукариот может происходить сразу в нескольких точках одной молекулы ДНК; скорость у эукариот около 100, а у бактерий - около 1000 нуклеотидов в сек.
Репликон - единица процесса репликации участка генома, который находится под контролем одной точки инициации (начала) репликации. Термин предложен Ф. Жакобом и С. Бреннером в 1963 году. Геном прокариот представляет собой, как правило, один репликон. От точки инициации репликация идёт в обе стороны, в некоторых случаях с неравной скоростью. У эукариот геном состоит из многих (часто до неск. десятков тысяч) репликонов.
Генетический код, его свойства.
Генетический код – способ записи генетической инофрмации о последовательности аминокислот в белке с помощью нуклеотидов. Открытие ген. Кода принадлежит Георгию Гамову. 1954год.
Триплетность - значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Непрерывность - между триплетами нет знаков препинания, то есть информация считывается непрерывно.
Неперекрываемость - один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
Однозначность (специфичность) - определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты - цистеин и селеноцистеин)
Вырожденность (избыточность) - одной и той же аминокислоте может соответствовать несколько кодонов.
Универсальность - генетический код работает одинаково в организмах разного уровня сложности - от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
Помехоустойчивость - мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными ; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными .
Понятие о гене. Свойства Гена.
Ген - структурная и функциональная единица наследственности живых организмов. Ген представляет собой последовательность ДНК, задающую последовательность определённого полипептида либо функциональной РНК. Гены определяют наследственные признаки организмов, передающиеся от родителейпотомству при размножении. При этом некоторые органеллы (митохондрии, пластиды) имеют собственную ДНК, не входящую в геном организма, которая определяет их признаки.
(Термин введен в 1909 году датским ботаником Вильгельмом Йогансеном)
дискретность - несмешиваемость генов;
стабильность - способность сохранять структуру;
лабильность - способность многократно мутировать;
множественный аллелизм - многие гены существуют в популяции во множестве молекулярных форм;
аллельность - в генотипе диплоидных организмов только две формы гена;
специфичность - каждый ген кодирует свой признак;
плейотропия - множественный эффект гена;
экспрессивность - степень выраженности гена в признаке;
пенетрантность - частота проявления гена в фенотипе;
амплификация - увеличение количества копий гена.
Особенности организации генома эукариот.
Геном эукариот:
большое число генов,
большее количество ДНК,
в хромосомах имеется очень сложная система контроля активности генов во времени и пространстве, связанная с дифференциацией клеток и тканей в онтогенезе организма.
Количество ДНК в хромосомах велико и возрастает по мере усложнения организмов. Для эукариот также характернаизбыточность генов. Так, у человека геном содержит число нуклеотидных пар, достаточное для образования более 2 млн. структурных генов, в то время как у человека имеется по данным 2000 года 31 тыс. всех генов.
Больше половины гаплоидного набора генома эукариотов составляют уникальные гены, представленные лишь по одному разу. У человека таких уникальных генов - 64%, у теленка - 55%, у дрозофилы - 70%.
Классы нуклеотидных последовательностей в ДНК эукариот, их характеристика, свойства и биологичесок значение.
Нуклеотидные последовательности в геноме эукариот
В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1. Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (~ 10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина.
Уровни организации генома эукариот.
Химический и структурный состав хромосом.
Молекулярно-биологические исследования позволили получить представление не только о химической структуре хромосом, но также и об их надмолекулярной организации и особенностях функционирования. В настоящее время известно, что хромосомы представляют собой нуклеопротеидные образования, состоящие из ДНК и белка. Кроме того, в хромосомах присутствует некоторое количество РНК, образующейся при транскрипции, и ионы Са+ и Mg+. Каждая хроматида, а в промежутке времени анафаза- S -период интерфазы и хромосома, содержит одну молекулу ДНК, которая определяет все функции хромосомы, связанные с хранением наследственной информации, её передачей и реализацией. Молекула ДНК в хромосомах тесно связана с двумя классами белков- гистонами (основные белки) и негистонами (кислые белки). Гистоны - это небольшие по величине белки с высоким содержанием заряженных аминокислот (лизина и аргинина). Суммарный положительный заряд позволяет гистонам связываться с ДНК независимо от нуклеотидного состава. Им принадлежит в основном структурная функция. Это очень стабильные белки, молекулы которых могут сохраняться в течение всей жизни клетки. В эукариотической клетке присутствуют 5 типов гистонов, которые распределяются на две основные группы: первая группа (их обозначают как Н2А, Н2В, НЗ, Н4), отвечает за формирование специфических дезоксирибонуклеопротеидных комплексов - нуклеосом. Вторая группа гистонов (HI) располагается между нуклеосомами и фиксирует укладку нуклеосомной цепи в более высокий уровень структурной организации (супернуклеосомную нить). Среди гистоновых белков, кроме структурных, встречаются такие, которые способны ограничивать доступность ДНК для ДНК - связывающих регуляторных белков и тем самым участвовать в регуляции активности генов. Негистоновые белки весьма разнообразны. Число их фракций превышает 100. Они присутствуют в меньших количествах в хромосомах в сравнении с гистонами и выполняют в основном регуляторную функцию. Участвуют в регуляции транскрипционной активности генов, в обеспечении редупликации и репарации ДНК. Большинство негистоновых белков хроматина присутствуют в клетках в небольшом количестве (минорные) - это регуляторные белки, узнающие специфические последовательности ДНК и связывающиеся с ними. Они вовлечены во многие генетические процессы, но известно о них пока что немного. Количественно преобладают негистоновые белки (мажорные), высокоподвижные, относительно малого размера, с большим электрическим зарядом - они всегда соединяются с нуклеосомами, содержащими активные гены. Кроме того, в группу негистоновых белков входит много ферментов.
Уровни упаковки наследственного материала у эукариот.
Таким образом, уровни упаковки ДНК следующие:
1) Нуклеосомный (2,5 оборота двуспиральной ДНК вокруг восьми молекул гистоновых белков).
2) Супернуклеосомный - хроматиновая спираль (хромонема).
3) Хроматидный - спирализованная хромонема.
4) Хромосома - четвертая степень сперализации ДНК.
В интерфазном ядре хромосомы деконденсированы и представлены хроматином. Деспирализованный участок, содержащий гены, называется эухроматин (разрыхленный, волокнистый хроматин). Это необходимое условие для транскрипции. Во время покоя между делениями определенные участки хромосом и целые хромосомы остаются компактными.
Эти спирализованные, сильно окрашивающиеся участки, называются гетерохроматином. Они неактивны в отношении транскрипции. Различают факультативный и конститутивный гетерохроматин.
Факультативный гетерохроматин информативен, т.к. содержит гены и может переходить в эухроматин. Из двух гомологичных хромосом одна может гетерохроматической. Конститутивный гетерохроматин всегда гетерохроматичен, неиформативен (не содержит генов) и поэтому всегда неактивен в отношении транскрипции.
Хромосомная ДНК состоит из более 10 8 пар оснований, из которых образуется информативные блоки - гены, расположенные линейно. На их долю приходится до 25% ДНК. Ген - функциональная единица ДНК, содержащая информацию для синтеза полипептидов, или всех РНК. Между генами находятся спейсеры - неинформативные отрезки ДНК разной длины. Избыточные гены представлены большим числом - 10 4 идентичных копий. Примером являются гены для т-РНК, р-РНК, гистонов. В ДНК встречаются последовательности одних и тех же нуклеотидов. Они могут быть умеренно повторяющимися и высоко повторяющимися последовательностями. Умеренно повторяющиеся последовательности достигают 300 пар нуклеотидов с повторениями 10 2 - 10 4 и представляют чаще всего спейсеры, избыточные гены.
Высокоповторяющиеся последовательности (10 5 - 10 6) образуют конститутивный гетерохроматин. Около 75% всего хроматина не участвует в транскрипции, он приходится на высокоповторяющиеся последовательности и нетранскрибируемые спейсеры.
Морфологические особенности метафазной хромосомы.
Митотическая суперкомпактизация хроматина делает возможным изучение внешнего вида хромосом с помощью световой микроскопии. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры или кинетохора ) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
В зависимости от места положения центромеры и длины плеч, расположенных по обе стороны от нее, различают несколько форм хромосом: равноплечие, или метацентрические (с центромерой посередине), неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов), палочковидные, или акроцентрические (с центромерой, расположенной практически на конце хромосомы), и точковые -очень небольшие, форму которых трудно определить (рис. 3.52). При рутинных методах окраски хромосом они различаются по форме и соотносительным размерам. При использовании методик дифференциальной окраски выявляется неодинаковая флуоресценция или распределение красителя по длине хромосомы, строго специфические для каждой отдельной хромосомы и ее гомолога (рис. 3.53).
Таким образом, каждая хромосома индивидуальна не только по заключенному в ней набору генов, но и по морфологии и характеру дифференциального окрашивания.
Эу- и гетерохроматин, их биологическое значение.
Некоторые хромосомы во время клеточного деления выглядят конденсированными и интенсивно окрашенными. Такие различия были названы гетеропикнозом. Для обозначения районов хромосом, демонстрирующих положительный гетеропикноз на всех стадиях митотического цикла был предложен термин «гетерохроматин». Различают эухроматин - основную часть митотических хромосом, которая претерпевает обычный цикл компактизации декомпактизации во время митоза, и гетерохроматин - участки хромосом, постоянно находящиеся в компактном состоянии.
У большинства видов эукариот хромосомы содержат как эу-, так и гетерохроматиновые участки, причем последние составляют значительную часть генома. Гетерохроматин располагается в прицентромерных, иногда в прителомерных областях. Обнаружены гетерохроматиновые участки в эухроматиновых плечах хромосом. Они выглядят как вкрапления (интеркаляции) гетерохроматина в эухроматин. Такой гетерохроматин называют интеркалярным. Компактизация хроматина. Эухроматин и гетерохроматин различаются по циклам компактизации. Эухр. проходит полный цикл компактизации-декомпактизации от интерфазы до интерфазы, гетеро. сохраняет состояние относительной компактности. Дифференциальная окрашиваемость. Разные участки гетерохроматина окрашиваются разными красителями, некоторые районы - каким-то одним, другие - несколькими. Применяя различные красители и используя хромосомные перестройки, разрывающие гетерохроматиновые районы, у дрозофилы удалось охарактеризовать много небольших районов, где сродство к окраскам отлично от соседних участках.
Понятие о кариотипе (определение).Обща характеристика кариотипа человека.
Кариотип - диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определенным числом, строением и генетическим составом хромосом.
Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где значение п различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некоторыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом (аутосомы ), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы ). Иногда эти различия состоят в разном количестве гетерохромосом у самок и самцов (XX или ХО). Чаще различия касаются строения половых хромосом, обозначаемых разными буквами -X и Y (XX или XY).
Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключенный в кариотипе,- генотип - это уникальное сочетание парных аллелей генома. В генотипе содержится программа развития конкретной особи.
Денверская (1960) и Парижская (1971) классификация хромосом человека: основные принципы и сущность.
Денверская и Парижская классификация хромосом Хромосомы подразделяются на аутосомы (соматических клеток) и гетерохромосомы (половых клеток). По предложению Левитского (1924) диплоидный набор соматических хромосом клетки был назван кариотипом. Он характеризуется числом, формой, размерами хромосом. Для описания хромосом кариотипа по предложению С.Г. Навашина их располагают в виде идиограммы - систематизированного кариотипа. В 1960 году была предложена Денверская международная классификация хромосом, где хромосомы классифицированы по величине и расположению центромеры. В кариотипе соматической клетки человека различают 22 пары аутосом и пару половых хромосом. Набор хромосом в соматических клетках называют диплоидным , а в половых клетках - гаплоидным (он равен половине набора аутосом). В идиограмме кариотипа человека хромосомы делят на 7 групп, в зависимости от их размеров и формы. 1 - 1-3 крупные метацентрические. 2 - 4-5 крупные субметацентрические. 3 - 6-12 и Х-хромосома средние метацентрические. 4 - 13-15 средние акроцентрические. 5 - 16-18 относительно малые мета-субметацентрические. 6 - 19-20 малые метацентрические. 7 - 21-22 и Y-хромосома наиболее малые акроцентрические. Согласно Парижской классификации хромосомы разделены на группы по их размерам и форме, а также линейной дифференцировке.
- это сложные мономеры, из которых собраны гетерополимерные молекулы. ДНК и РНК. Свободные нуклеотиды участвуют в сигнальных и энергетических процессах жизнедеятельности. ДНК-нуклеотиды и РНК-нуклеотиды имеют общий план строения, но различаются по строению сахара-пентозы. В ДНК-нуклеотидах используется сахар дезоксирибоза, а в РНК-нуклеотидах - рибоза.
Структура нуклеотида
В каждом нуклеотиде можно выделить 3 части:
1. Углевод - это пятичленный сахар-пентоза (рибоза или дезоксирибоза).
2. Фосфорный остаток (фосфат) - это остаток фосфорной кислоты.
3. Азотистое основание - это соединение, в котором много атомов азота. В нуклеиновых кислотах используется всего 5 видов азотистых оснований: Аденин, Тимин, Гуанин, Цитозин, Урацил. В ДНК - 4 вида: Аденин, Тимин, Гуанин, Цитозин. В РНК - тоже 4 вида: Аденин, Урацил, Гуанин, Цитозин, Легко заметить, что в РНК происходит замещение Тимина на Урацил по сравнению с ДНК.
Общая структурная формула пентозы (рибозы или дезоксирибозы), молекулы которой образуют "скелет" нуклеиновых кислот:
Если Х заменить на Н (Х = Н) - то получаются дезоксирибонуклеозиды; если Х заменить на ОН (Х = ОН) - то получаются рибонуклеозиды. Если вместо R подставить азотистое основание (пуриновое или пиримидиновое) - то получится конкретный нуклеотид.
Важно обратить внимание на те положения атомов углерода в пентозе, которые обозначены как 3" и 5". Нумерация атомов углерода начинается от атома кислорода вверху и идёт по часовой стрелке. Последним получается атом углерода (5"), который располагается за пределами пентозного кольца и образует, можно сказать, "хвостик" у пентозы. Так вот, при наращивании цепочки из нуклеотидов фермент может присоединить новый нуклеотид только к углероду 3" и ни к какому другому. Поэтому 5"-конец нуклеотидной цепочки никогда не сможет иметь продолжения, удлинняться может только 3"-конец.


Сравните нуклеотид для РНК с нуклеотидом для ДНК.
Попробуйте узнать, какой это нуклеотид, в таком представлении:

АТФ - свободный нуклеотид

цАМФ - "закольцованная" молекула АТФ
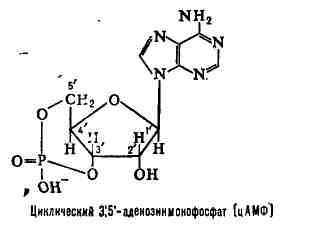
Схема строения нуклеотида

Обратите внимание на то, что активированный нуклеотид, способный наращивать цепочку ДНК или РНК, имеет "трифосфатный хвостик". Именно этим "энергонасыщенным" хвостиком он может присоединиться к уже имеющейся цепочке растущей нуклеиновой кислоты. Фосфатный хвостик сидит на 5-м атоме углерода, так что это положение углерода уже занято фосфатами и предназнено для прикрепления. К чему же его прикрепить? Только к углероду в положении 3". После прикрепления данный нуклеотид сам станет мишенью дла прикрепления следующего нуклеотида. "Принимающая сторона" предоставляет углерод в положении 3", а "прибывающая сторона" цепляется к нему фосфатным хвостиком, находящимся в положении 5". В целом цепочка растёт со стороны 3".
Наращивание нуклеотидной цепочки ДНК
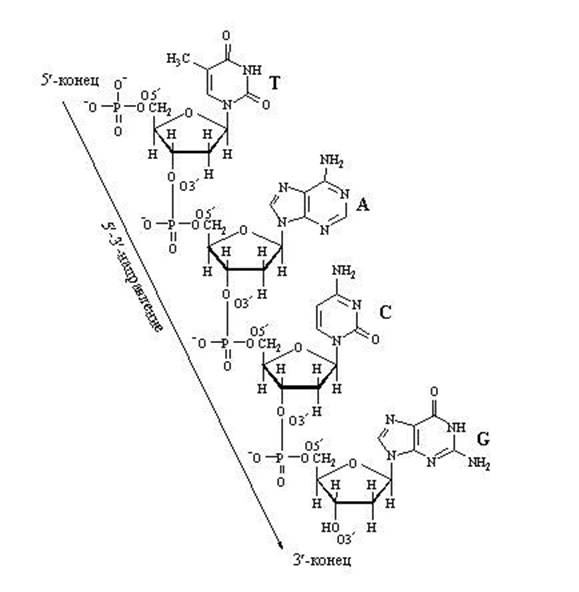
Наращивание цепочки за счёт "продольных" связей между нуклеотидами может идти только в одном направлении: от 5" ⇒ к 3", т.к. новый нуклеотид можно присоединить только к 3"-концу цепочки, но не к 5"-концу.
Пары нуклеотидов, связанные "поперечными" комплементарными связями своих азотистых оснований
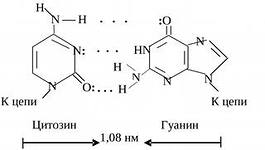
Участок двойной спирали ДНК

Найдите признаки антипараллельности двух цепей ДНК.
Найдите пары нуклеотидов с двойными и тройными комплементарными связями.










