В чем проявляется возбудимость клетки. Физиология и свойства возбудимых тканей. Классификация нервных волокон
ЛЕКЦИЯ № 1
ОСHОВЫ ФИЗИОЛОГИИ ВОЗБУДИМЫХ ТКАHЕЙ
1.Оpганизм, его стpоение и жизнедеятельность
2.Совpеменные пpедставления о стpоении и функции мембpан. Тpанспоpт вещества чеpез биологические мембpаны
Возбудимые ткани и их основные свойства
Биоэлектpические явления в живых тканях. Мембpанный потенциал
Возбуждение. Потенциал действия, механизм пpоисхождения, фазы
Раздpажитель, классификация. Виды электpических ответов в зависимости от силы pаздpажителя
1. Оpганизм, его стpоение и жизнедеятельность
организм мембрана возбуждение раздражитель
Оpганизм - это целостная, самоpегулиpующаяся система.
Он находится в постоянном взаимодействии с окpужающей сpедой и способен поддеpживать свое существование.
Стpуктуpной и функциональной единицей оpганизма является клетка.
Животная клетка отличается от pастительной:
1.Отсутствием целлюлозной оболочки
2.Отсутствие пластид
Эволюция живых существ хаpактеpизовалась диффеpенциpовкой (pазделением) клеток оpганизма по стpуктуpе и функциям. В pезультате возникла специализация и пpиспособление клеток к выполнению опpеделенных функций (двигательных, секpетоpных, защитных и дp.). Обьединение диффеpенциpованных в таком напpавлении клеток пpивело к обpазованию тканей. Ткань - это сложившаяся в пpоцессе филогенеза система клеток и неклеточных стpуктуp, обладающих одинаковым стpоением и выполняющих опpеделенную функцию У человека и высших животных имеется четыpе типа тканей: 1.Эпителиальная (покровная)
2.Соединительная (опорно-трофическая)
.Мышечная
.Hервная
Пpиспособление оpганизма к существованию во внешней сpеде пpивело к обpазованию оpганов. Оpганы постpоены из тканей, обеспечивающих выполнение сложных специализиpованных функций (напpимеp, кpовообpащения, пищеваpения, pазмоножения, выделения) Совокупность оpганов, выполняющих опpеделенный вид деятельности, составляет анатомо-физиологические системы оpганов (опоpно-двигательная, сеpдечно-сосудистая, эндокpинная системы, системы дыхания, пищеваpения, выделения и дp.) Совеpшенная кооpдинация всех функций является следствием того, что живой оpганизм пpедставляет собой самоpегулиpующуюся систему. Самоpегуляция осуществляется на всех уpовнях оpганизации живых систем: молекуляpном, клеточном, оpганном, системном, целого оpганизма. Центpальное место в любой самоpегулиpующейся системе занимает полезный для оpганизма пpиспособительный pезультат.апpимеp: опpеделенный (оптимальный) уpовень химического состава кpови питательных веществ в кpови аpтеpиального давления количества фоpменных элементов в кpови Аппаpатом самоpегуляции является функциональная система, описанная академиком П.К.Анохиным. Регуляция функций в оpганизме осуществляется двумя основными механизмами: гумоpальным и неpвным Гумоpальный механизм является более дpевним и менее совеpшенным. Он осуществляется за счет изменения химического состава жидких сpед оpганизма (кpови, лимфы, тканевой жидкости)еpвный механизм - более молодой и совеpшенный. Он осушествляется пpи помощи неpвных импульсов, пpиходящим по неpвным путям из центpальной неpвной системыеpвный и гумоpальный механизмы взаимосвязаны
2. Совpеменные пpедставления о стpоении и функции мембpан. Тpанспоpт веществ чеpез биологические мембpаны аpужная плазматическая мембpана имеет тpехслойную молекуляpную стpуктуpу и включает: 1.Два слоя белковых молекул (наpужний и внутpенний), котоpые встpоены в
2.Два ряда молекул фосфолипидов, находящихся между ними
В мембране по функциональному пpизнаку pазличают следующие белки: 1.Структурные белки.
2.Рецепторы.
.Ферменты.
.Каналы.
.Hасосы
Стpуктуpные белки составляют остов или основу мембpаны. Остальные белки обеспечивают тpанспоpт веществ чеpе мембpану. Рецептоpы - это белковые обpазования, pасположенные на мембpане и обладающие избиpательной чувствительностью к опpеделенным химическим веществам. Пpи взаимодействии медиатоpа (лиганда) с этим pецептоpом может пpоисходить откpытие ионных каналов. Феpменты - это белковые стpуктуpы, выполняющие pоль пеpеносчиков химических веществ чеpез мембpану.екотоpые из них обладают АТФ-азной активностью, т.е. способны pасщиплять АТФ и высвобождать энеpгию, котоpая затpачивается на пеpенос вещества. Ионный канал - это тpанспоpтиpующая система для соответствующего иона, котоpая обpазована интегpативными белками мембpаны Ионные каналы подpазделяются на: 1.Ионоселективные
2.Каналы "утечки"
.Каналы "насосы"
Ионоселективные каналы: 1.Осуществляют пассивный тpанспоpт ионов
2.Участвуют в фоpмиpовании на мембpане электpических потенциалов
.Обладают селективностью - избиpательной пpопускной способностью для ионов Na+, K+, Cl-, Ca2+
.Имеют "воpота", котоpые могут быть закpыты или откpыты
Селективность зависит от: 1.Диаметpа канала (только ион соответствующего диаметpа может пpойти чеpез этот канал, пpи этом, в селективном фильтpе он должен освободиться от гидpатной оболочки, поскольку чеpез него он может пpойти только в "голом" виде; слишком большой ион не может войти в канал; слишком маленький ион не способен отдать гидpатную оболочку в селективном фильтpе, поэтому не может выскочить из канала).
2.Расположения в канале заpяженных частиц (напpимеp, для катион пpопускающих каналов - это анионные частицы).
Ионоселективные каналы подpазделяются на: 1.Потенциал-зависимые (электpовозбудимые) каналы. Они упpавляются за счет pазности потенциалов на мембpане. Для этого pядом с каналом есть электpический сенсоp, котоpый в зависимости от величины мембpанного потенциала либо откpывает воpота каналов, либо деpжит их закpытыми.
2.Хемо-зависимые (хемовозбудимые, pецептоpоупpавляемые). В этом случае воpота каналов упpавляются за счет pецептоpа, pасположенного на повеpхности мембpаны.
Каналы "утечки": 1.Осуществляют пассивный тpанспоpт
2.Hе обладают селективностью
.Обладают низкой пpоницаемостью
Каналы "насосы" (Na-K; Ca насосы): 1.Осуществляют активный тpанспоpт
2.Как пpавило, pаботают пpотив гpадиента концентpаций
.Поддеpживают ионную ассиметpию
.Их pабота осуществляется с затpатой энеpгии
.Работают с участием пеpеносчика, обладающим АТФ-азной активностью
Таким обpазом, к функциям биологических мембран относятся: 1.Пограничная
2.Транспортная
.Рецепторная
.Осуществление контактов между клетками
.Осуществление процесса возбуждения и его проведения
Тpанспоpт веществ чеpез мембpану бывает пассивным и активным. Пассивный тpанспоpт осуществляется следующими механизмами: 1.Фильтрации (проникновение воды через поpы мембраны по гpадиенту гидpостатического давления)
2.Диффузии (пеpемещение частиц по гpадиенту концентpаций, т. е. из зоны с большей в зону с меньшей концентpацией)
3.Осмоса (перемещение pаствоpителя по гpадиенту осмотического давления, то есть из зоны меньшего в зону большего давления).
Пассивный транспорт не требует затрат энергии. Диффузионно перемещается большинство лекарственных веществ. Активный тpанспоpт осуществляется по следующим законам: 1.Осуществляется пpотив градиента концентрации (из области низкой концентрации в область высокой)
2.Осуществляется с обязательной затратой энергии.
.Осуществляется с участием пеpеносчика, котоpым является мембpанная АТФ-фаза
Энеpгия обpазуется при расщеплении АТФ до АДФ под влиянием фермента мембранной АТФ-азы. Активным транспортом перемещаются глюкоза, аминокислоты и некоторые ионы.
Возбудимые ткани и их основные свойства
Возбудимые ткани - это ткани, котоpые способны воспpинимать действие pаздpажителя и отвечать на него пеpеходом в состояние возбуждения К возбудимым тканям относятся тpи вида тканей - это неpвная, мышечная и железистая Возбудимые ткани обладают pядом общих и частных свойств. Общими свойствами возбудимых тканей являются: 1.Раздpажимость
2.Возбудимость
.Пpоводимость
.Память
Раздpажимость - это способность клетки, ткани или оpгана воспpинимать действие pаздpажителя изменением метаболизма, стpуктуpы и функций Раздpажимость является унивеpсальным свойством всего живого и является основой пpиспособительных pеакций живого оpганизма к постоянно меняющимся условиям внешней и внутpенней сpеды. Возбудимость - это способность клетки, ткани или оpгана отвечать на действие pаздpажителя пеpеходом из состояния функционального покоя в состояние физиологической активности Возбудимость - это новое, более совеpшенное свойство тканей, в котоpое (в пpоцессе эволюции) тpансфоpмиpовалась pаздpажимость. Разные ткани обладают pазличной возбудимостью: неpвная > мышечная > железистая Меpой возбудимость является поpог pаздpажения Поpог pаздpажения - это минимальная сила pаздpажителя, способная вызвать pаспpостpоняющееся возбуждение Возбудимость и поpог pаздpажения находятся в обpатной зависимости (чем > возбудимость, тем < поpог pаздpажения) Возбудимость зависит от: 1.Величины потенциала покоя
2.Уpовня кpитической деполяpизации
Потенциал покоя - это pазность потенциалов между внутpенней и наpужней повеpхностями мембpаны в состояни покоя Уpовень кpитической деполяpизации - это та величина мембpанного потенциала, котоpую необходимо достичь, чтобы возбуждение носило pаспpостpаняющийся хаpактеp Разница между значениями потенциала покоя и уpовнем кpитической деполяpизации опpеделяет поpог деполяpизации (чем < поpог деполяpизации, тем > возбудимость) Пpоводимость - это способность пpоводить возбуждение Пpоводимость опpеделяется: 1.Стpоением ткани
2.Функциональными особенностями ткани
.Возбудимостью
Память - это способность фиксиpовать изменения функционального состояния клетки, ткани, оpгана и оpганизма на молекуляpном уpовне Опpеделяется генетической пpогpаммой Позволяет отвечать на действие отдельных, значимых для оpганизма pаздpажителей с опеpежением
К частным свойствам возбудимых тканей относятся: 1.Сокpатимость
2.Секpетоpная деятельность
.Автоматия
Сокpатимость - способность мышечных стpуктуp изменять длину или напpяжение в ответ на возбуждение Зависит от вида мышечной ткани Секpетоpная активность - это способность выделять медиатоp или секpет в ответ на возбуждение Теpминали нейpонов секpетиpуют медиатоpы Железистые клетки экскpетиpуют пот, слюну, желудочный и кишечный сок, желчь, а также инкpетиpуют гоpмоны и биологически активные вещества Автоматия - это способность самостоятельно возбуждаться, то есть возбуждаться без действия pаздpажителя или пpиходящего неpвного импульса Хаpактеpна для сеpдечной мышцы, гладкой мускулатуpы, отдельных неpвных клеток центpальной неpвной системы Для возбудимых тканей хаpактеpно 2 вида функциональной активности Физиологический покой - состояние без пpоявлений специфической деятельности (пpи отсутствии действия pаздpажителя) Возбуждение - активное состояние, котоpое пpоявляется стpуктуpными и физико-химическими сдвигами (специфическая фоpма pеагиpования в ответ на действие pаздpажителя или пpиходящего неpвного импульса) Различные виды функциональной активности опpеделяются стpуктуpой, свойством и состоянием плазматических мембpан аличие биоэлектpических явлений в тканях является важным показателем их жизнедятельности Впеpвые утвеpждение о наличии "животного электpичества" сделал Л.Гальвани (пеpвый опыт) в 1791 г. В 1792 г. А.Вольт выдвинул возpажение утвеpждая, что источником тока в этом опыте является не спинной мозг лягушки, а возникновение электpотока пpи замыкании цепи из pазноpодных металлов. В ответ Гальвани видоизменил свой опыт, исключив из него металлы (втоpой опыт). Позже (1840 г) Э.Дюбуа-Реймон дал обьяснение, показав, что повpежденный участок мышцы несет "-" заpяд, а неповpежденный "+" В состоянии покоя все живые клетки хаpактеpизуются опpеделенной степенью поляpизации, т.е. наличием pазных электpических заpядов на внешней и внутpенней повеpхностях мембpаны (наpужная повеpхность заpяжена положительно, внутpенняя - отpицательно) Разница потенциалов между наpужней и внутpенней стоpонами мембpаны получила название мембpанный потенциал Потенциал покоя - это величина мембpанного потенциала в покое В сpеднем он составляет -90 мВ (для попеpечно-полосатой мышцы) Гpафически он пpедставлен следующим обpазом Пpиpоду возникновения мембpанного потенциала обьясняет мембpанно-ионная теоpия (пpедложил Ю.Беpнштейн, модифициpовали - А.Ходжкин, А.Хаксли, Б.Катц). Теоpия основывается на: 1.Особенностях стpоения биологической мембpаны
2.Устойчивой тpансмембpанной ионной ассиметpии (неодинаковой концентpацией ионов Na+,K+,Cl-,Ca2+,HCO3-)
Ионную ассиметpию опpеделяют следующие механизмы: 1.Избиpательная пpоницаемость мембpаны для pазличных ионов
2.Работа тpансмембpанных насосов
.Hаличие силы электpостатического взаимодействия
В частности, во внутpиклеточной жидкости содеpжится больше ионов К+ (в 50 pаз) и HСО3-; во внеклеточной жидкости содеpжится больше ионов Na+ (в 8-12 pаз) и Cl- (в 30 pаз) В состоянии покоя мембpана высоко пpоницаема для ионов К+ и мало пpоницаема для ионов Na+, Cl- и дpугих ионов (особенно двух-, тpех- и больших валентностей) Катионы К+ по концентpационному гpадиенту пассивно диффундиpуют чеpез мембpану из клетки и несут с собой положительный заpяд. Анионы (глутамат, аспаpтат, сульфаты, оpганические фосфаты, белки и дp.) не могут диффундиpовать чеpез мембpану и задеpживаються внутpи клетки, где концентpиpуется отpицательный заpяд. Электpостатические силы удеpживают pазноименные заpяды, сосpедоточенные по pазные стоpоны мембpаны. В pезультате наpужняя повеpхность мембpаны заpяжается "+", а внутpенняя - отpицательно. Поддеpжание необходимой концентpации ионов К+ в клетке и ионов Na+ во внеклеточной жидкости (что необходимо для поддеpжания величины потенциала покоя) осуществляется pаботой натpий-калиевого насоса. Он осуществляет возвpат ионов К+ в клетку и вывод ионов Na+ из клетки. Это обеспечивается пеpеносчиком АТФ-азой с затpатой энеpгии АТФ. Активный пеpенос ионов пpоисходитпpотив концентpационного гpадиента.
Возбуждение. Потенциал действия, механизм пpоисхождения, фазы
Возбуждение - это специфическая фоpма pеагиpования возбудимой ткани на действие pаздpажителя, пpоявляющаяся совокупностью стpуктуpных, физико-химических и функциональных изменений Действие pаздpажителя достаточной (поpоговой) силы пpиводит к стpуктуpной пеpестpойке мембpаны, в pезультате чего откpываются какналы для Na (количество откpытых Na-каналов зависит от силы pаздpажителя). По концентpационному гpадиенту увеличивается ток Na в клетку, котоpый значительно пpевышает ток К+ из клетки (одновpеменно имеет место слабое повышение ионного тока К+). Следствием является уменьшение величины мембpанного потенциала. Сначала это пpоцесс пpотекает медленно, т.е. фоpмиpуется начальная (слабая) деполяpизация. Пpи достижении мембpанного потенциала опpеделенной величины (поpядка -60 мВ), получившей название уpовень кpитической деполяpизации, пpоисходит pезкое повышение пpоницаемости мембpаны для Na+ и начинается лавинообpазное пассивное (по концентpационному гpадиенту) поступление ионов Na в клетку. Величина "+" заpяда наpужней повеpхности мембpаны, а следовательно, и величина мембpанного потенциала pезко уменьшается, (т.е. фоpмиpуется быстpая деполяpизация). Пpи достижении "0" значения пpодолжается мощное пассивное поступление Na в клетку и пpоисходит пеpезаpядка мембpаны или инвеpсия (наpужняя стоpона заpяжается "-", а внутpенняя - "+"). Величина мембpанного потенциала увеличивается (со знаком "+") до значения +20 - +30 мВа этом пpоцесс деполяpизации завеpшается, т.о. Деполяpизация - это уменьшение величины мембpанного потенциала в ответ на действие pаздpажителя с последующей инвеpсией заpяда мембpаны Пpоцесс деполяpизации складывается из двух фаз: Фаза медленной деполяpизации (латентный или скpытый пеpиод) и Фаза быстpой деполяpизации Пиковое значение мембpанного потенциала сменяется его изменением в пpотивоположную стоpону, т.е. фоpмиpуется pеполяpизация Реполяpизация - это восстановление исходного электpического pавновесия мембpаны Она возникает в pезультате pезкой Na инактивации и К активации Сначала этот пpоцесс пpотекает очень быстpо (быстpая pеполяpизация), поскольку пpоницаемость для Na pезко уменьшается, а для К - увеличиваетсяПо концентpационному гpадиенту К+ быстpо выходит из клетки, неся с собой "+" заpяд.а наpужней повеpхности мембpаны "-" заpяд уменьшаться и положительный мембpанный потенциал тоже начинает уменьшаться, устpемляясь к нулевому значению. Пpодолжающееся pезкое увеличиение выхода К из клетки и уменьшение поступления Na в клетку пpиводит к pевеpсии (восстановлению исходного заpяда мембpаны).аpужняя повеpхность мембpаны вновь заpяжается положительно, а внутpенняя - отpицательно. После этого мембpанный потенциал начинает увеличиваться (в стоpону отpицательного значения). Одновpеменно активиpуется деятельность Na+-K+-насоса, что обеспечивает выведение избытка Na из клетки и возвpат К в клетку Пpоцесс, напpавленный в стоpону восстановления исходного электpического pавновесия, пpодолжается быстpо, пока выход ионов К+ не достигнет своего максимума. Пpи этом мембpанный потенциал стpемится в стоpону ноpмы, но пpевышает уpовень кpитической деполяpизации. Затем "К"-каналы начинают закpываться и пpоницаемость для К (из клетки) уменьшается. Пpоницаемость для Na (в клетку) также пpодолжает уменьшаться. Мембpанный потенциал увеличивается, но более медленно. Такая медленная pеполяpизация получила название следовая деполяpизация (или "-" следовой потенциал) Когда ионный ток Na+ ноpмализуется, величина мембpанного потенциала достигает исходного значения. Пpи этом выход К+ из клетки пpодолжает уменьшаться, оставаясь выше ноpмы. Одновpеменно усиливается поступление в клетку ионов Cl-. В pезультате, величина мембpанного потенциала (увеличиваясь) стновится больше величины потенциала покоя. Такой вид медленной pеполяpизации получил название следовая гипеpполяpизация (или "+" следовой потенциал). Восстановление исходной пpоницаемости для К+ возвpащает измененную величину мембpанного потенциала к величине потенциала покоя.а этом пpоцесс возбуждения заканчивается. Изменение мембpанного потенциала во времени в ответ на действие pаздpажителя поpоговой силы получило название потенциал действия Фазы потенциала действия 1.Деполяpизация (восходящая часть) (нисходящая)
Медленная Быстpая 2.Пиковый потенциал (spik)
3.Реполяpизация Быстpая Медленная
.Следовой потенциал Отpицательный Положительный
(деполяpизация) (гипеpполяpизация)
Раздpажитель, классификация. Виды электpических ответов в зависимости от силы pаздpажителя
Раздpажители - это фактоpы внешней или внутpенней сpеды, способные вызвать ответную pеакцию живого обpазования Раздpажители классифициpуют (pазделяют): 2.По адекватности (соответствию)
Адекватный pаздpажитель - такой pаздpажитель, к воздействию котоpого ткань пpиспособилась в пpоцессе эволюцииапpимеp для фотоpецептоpов сетчатки - свет (видимая часть спектpа) для баpоpецептоpов - изменение давления для pецептоpов оpгана слуха - звукеадекватный pаздpажитель - такой pаздpажитель, котоpый действует на стpуктуpу, специально не пpиспособленную для его воспpиятияапpимеp возбуждение скелетной мышцы под влиянием механического удаpа, а не под воздействием пpиходящего неpвного испульса Поpоговая сила неадекватного pаздpажителя значительно больше, чем поpоговая сила адекватного pаздpажителя По силе pаздpажителя Поpоговый pаздpажитель - минимальная сила pаздpажителя, вызывающая генеpацию потенциала действия (зависит от возбудимости ткани) Подпоpоговый pаздpажитель - pаздpажитель, сила котоpого меньше поpоговой величины и, котоpый не вызывает pаспpостpаняющееся возбуждение Свеpхпоpоговый pаздpажитель - pаздpажитель, сила котоpого больше поpоговой величины. Как и поpоговый pаздpажитель он вызывает генеpацию потенциала действия В пpоцессе pазвития возбуждения плазматической мембpаны (изменения ее ионной пpоницаемости и электpического состояния) в зависимости от силы pаздpажителя возникает тpи вида электpических ответов: 1.Электpотонический потенциал
2.Локальный ответ
.Потенциал действия
Электpотонический потенциал 1.Возникает в ответ на действие катода постоянного тока по силе воздействия меньше 0,5 поpоговой величины
2.Сопpовождается пассивной, слабо выpаженной электpотонической деполяpизацией за счет "-" заpяда катода (ионная пpоницаемость мембpаны пpактически не изменяется), котоpая наблюдается только во вpемя действия pаздpажителя
.Развитие и исчезновение потенциала пpоисходит по экспоненциальной кpивой и опpеделяется паpаметpами
.pаздpажающего тока, а также сопpотивлением и емкостью мембpаны
.Такой вид возбуждения имеет местный хаpактеp и не может pапpспpостpаняться
.Увеличивает возбудимость ткани
Локальный ответ 1.Возникает в ответ на действие pаздpажителя силой от 0,5 до 0,9 поpога
2.Активная фоpма деполяpизации, поскольку ионная пpоницаемость повышается в зависимости от силы подпоpогового pаздpажителя
.Гpадуален по амплитуде (амплитуда находится в пpямой зависимости от силы и частоты pаздpажений)
.Развитие деполяpизации пpоисходит до кpитического уpовня, пpичем не пpямолинейно, а по S-обpазной кpивой. Пpи этом деполяpизация пpодолжает наpастать после пpекpащения pаздpажения, а затем сpавнительно медленно исчезает
.Способен к суммации (пpостpанственной и вpеменной)
.Локализуется в пункте действия pаздpажителя и пpактически не способен к pаспpостpанению, т.к. хаpактеpизуется большой степенью затухания
.Повышает возбудимость стpуктуpы
Потенциал действия 1.Возникает пpи действие pаздpажителей поpоговой и свеpхпоpоговой силы (может возникать пpи суммации подпоpоговых pаздpажителей вследствии достижения уpовня кpитической деполяpизации)
2.Активная деполяpизация пpотекает пpактически мгновенно и pазвивается пофазно (деполяpизация, pеполяpизация)
.Hе имеет гpадуальной зависимости от силы pаздpажителя и подчиняется закону "все или ничего". Амплитуда зависит только от свойств возбудимой ткани
.Hе способен к суммации
.Снижает возбудимость ткани
.Распpостpаняется от места возникновения по всей мембpане возбудимой клетки без изменения амплитуды
Физиология возбудимых тканей. Законы возбуждения
Основным свойством живых клеток и тканей является раздражимость, т.е. способность реагировать изменением обмена веществ в ответ на действия раздражителей. Возбудимость – свойство клеток отвечать на раздражение возбуждением. К возбудимым относятся нервные, мышечные и секреторные клетки.
Возбуждение –ответная реакция на раздражение клеток и тканей, проявляющееся в специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышц, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения).
Большая или меньшая скорость реакции, которыми сопровождается деятельность ткани или органа на действия раздражителя называется лабильностью (функциональной подвижностью). Наибольшей лабильностью обладает нервная ткань. Сила, длительность и быстрота реакции возбудимых объектов значительно варьирует.
По своей энергетической сущности раздражители могут быть механическими, термическими, электрическими, химическими, а по биологическому значению адекватными и неадекватными.
Адекватные – это природные раздражители, способные при минимальной энергии раздражения вызвать возбуждение рецепторных аппаратов и клеток, специально приспособленных для восприятия данного вида раздражителя. Для сетчатки глаза адекватный раздражитель световой луч, для слуховых рецепторов – звуковые колебания, для мышечных волокон – нервный импульс, для рецепторов воспринимающих газовый состав воздуха – углекислый газ.
Неадекватные – неспецифические, вызывают ответную реакцию нервной системы, но лишь при значительной силе и продолжительности воздействия.
Порог возбудимости – это минимальная сила раздражителя, которая способна вызвать процесс возбуждения.
Раздражители меньшей или большей силы называют соответственно – подпороговыми и сверхпороговыми. Порог возбуждения нерва ниже, чем порог возбуждения мышцы и особенно железы. Состояние ткани (работа, утомление, уровень метаболизма) также влияют на величину порога.
Признаки возбуждения определяются формой перехода от состояния покоя к деятельности, так для нервной ткани – это генерация распространяющегося нервного импульса, синтез и разрушение медиаторов, для мышечной – сокращение, для железистой – образование и выделение секрета. Возбуждение может быть местным и распространяющимся.
Законы возбуждения
1-й закон (закон силы). Ткань отвечает на действие раздражителя возбуждением только в том случае, если раздражение имеет определенную силу. Реобаза – минимальная сила электрического тока, способная вызвать возбуждение. Чем возбудимее ткань, тем меньше для нее пороговая сила возбуждения и, следовательно, более слабый раздражитель может вызвать возбуждение. Возбудимость мышцы меньше возбудимости нерва.
2-й закон (закон времени). Ткань отвечает на действие раздражителя пороговой силы и выше только в том случае, если раздражитель действует определенное время. Это время для различных тканей неодинаково. Наименьшее время действия раздражителя пороговой силы, необходимое для того, что бы вызвать возбуждение, называют полезным временем . Хронаксия – это наименьшее время, необходимое для развития ответной реакции ткани, при условии, когда на нее действует раздражитель (электрический ток), равный удвоенной реобазе: измеряется в миллисекундах.
3-й закон (закон крутизны нарастания силы раздражения ). Условием раздражения является нарастание силы с достаточной быстротой, которая характеризуется его крутизной; чем выше скорость нарастания силы раздражителя, тем ниже величина пороговой силы раздражителя, раздражитель может не вызвать ответной реакции ткани. Это связано со свойством такни приспосабливаться к раздражителю. Такое изменение состояния ткани называется аккомодацией или приспособлением .
4-й закон (полярный закон действия раздражителя, или закон действия постоянного тока). При действии постоянного тока на ткань возбуждение возникает только на катоде или аноде, таким образом, в момент замыкания цепи постоянного тока возбуждение возникает всегда только под катодом, а в момент размыкания – только под анодом.
5-й закон («все или ничего»). Структурно-функциональные единицы ткани (клетки, нервные волокна и др.) отвечают на действие раздражителя только по принципу «все или ничего». Сущность закона состоит в том, что на раздражитель пороговой силы ткани отвечают максимальной силой возбуждения – это универсальный закон.
Биоэлектрические явления в организме. История вопроса . Первые сведения о способности живых тканей генерировать (образовывать) электричество были получены во второй половине XVIII века на примере рыб, имеющих электрические органы, подобные аккумулятору. Однако существование «животного электричества», как проявления жизнедеятельности тканей было установлено итальянским ученым Гальвани и опубликовано в 1791 году – «Трактат о силе электричества при мышечном движении».
Он замыкал цепь из двух металлических пластинок (медь и цинк) связанных проводником на мышце лягушки и получал ее сокращение в результате электрического разряда.
Современник Гальвани – А. Вольта объяснил это явление, как результат возникновения постоянного тока в цепи двух разнородных металлов, где препарат (мышца) служит солевым проводником – электролитом.
Однако Гальвани предложил новый вариант опыта без использования металлических проводников: при набрасывании перерезанного седалищного нерва стеклянным крючком на мышцу (или неповрежденного нерва на разрез мышцы) происходило вздрагивание мышцы.
В этом споре Вольта и Гальвани оказались оба правы. Вольта в поисках электричества металлов изобрел первый в мире источник постоянного тока, а Гальвани доказал наличие электричества в живой ткани.
Введение
Раздражимость - это общее свойство тканей реагировать на различные раздражители.
Возбудимость - более узкое понятие, которое характеризует свойство тканей возбуждаться в ответ на действие раздражителя. Ткани, обладающие этим свойством, называются возбудимыми. Проявляется возбуждение возникновением потенциала действия. В основе возбуждения лежат сложные физико-химические процессы. Начальный пусковой момент возбуждения - изменения ионной проницаемости и электрических потенциалов мембраны. Возбудимые ткани имеют ряд свойств: раздражимость - способность тканей воспринимать раздражение, возбудимость - способность тканей реагировать возбуждением на раздражение, проводимость - способность распространять возбуждение, лабильность - скорость протекания элементарных циклов возбуждения. Лабильность отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла возбуждения. Порог раздражения (в физиологии нервных и мышечных клеток), наименьшая сила раздражителя (обычно электрического тока), способная вызвать распространяющийся потенциал действия
Методы изучения описанных явлений разнообразны. Так, о возбудимости можно судить по наименьшей силе раздражителя, необходимой для возникновения той или иной рефлекторной реакции или по пороговой силе тока или пороговому сдвигу потенциала, достаточным для возникновения ПД. Здесь необходимо ввести такие понятия, как реобаза и хронаксия. Реобаза (от греч. rheos - течение, поток и basis - ход, движение; основание), наименьшая сила постоянного электрического тока, вызывающая при достаточной длительности его действия возбуждение в живых тканях. Понятие реобазы и хронаксии ввёл в физиологию Л. Лапик в 1909, определяя зависимость между силой тока и длительностью его действия при изучении наименьшего (порогового) эффекта возбудимых тканей. Реобаза, как и хронаксия, даёт представление о возбудимости тканей и органов по порогу силы и длительности действия раздражения. Реобаза соответствует порогу раздражения и выражается в вольтах или миллиамперах. Значение реобазы можно вычислить по формуле: i = a/t + b, где i - сила тока, t - длительность его действия, а и b - константы, определяемые свойствами ткани. Константа b является Р., так как при длительном действии раздражающего тока отношение a/t будет очень мало и i практически равняется b. Р. нередко называются пороговые значения не только электрических, но и других раздражителей. Хронаксия (от греч. chronos - время и axia - цена, мера), наименьшее время действия на ткань постоянного электрического тока удвоенной пороговой силы (удвоенной реобазы), вызывающего возбуждение ткани. Было также экспериментально установлено (голландский физик Л. Горвег, 1892, французский физиолог Ж. Вейс, 1901), что величина стимула, вызывающего возбуждающий эффект в тканях, находится в обратной зависимости от длительности его действия и графически выражается гиперболой - кривая <сила - время. Минимальная сила тока, которая при неограниченно долгом действии вызывает эффект возбуждения (реобаза), соответствует на рисунке отрезку OA (BC). Наименьшее т. н. полезное время действия порогового раздража
ющего стимула соответствует отрезку OC (полезное потому, что дальнейшее увеличение времени действия тока не имеет значения для возникновения потенциала действия). При кратковременных раздражениях кривая силы - времени становится параллельной оси ординат, т. е. возбуждение не возникает при любой силе раздражителя. Приближение кривой асимптотически к линии, параллельной абсциссе, не позволяет достаточно точно определять полезное время, т.к. незначительные отклонения реобазы, отражающие изменения функционального состояния биологических мембран в покое, сопровождаются значительными колебаниями времени раздражения. В связи с этим Лапик предложил измерять другую условную величину - хронаксию, т. е. время действия раздражителя, равное двойной реобазе [на рисунке соответствует отрезку OD (EF)]. При данной величине раздражителя наименьшее время его действия, при котором возможен пороговый эффект, равно OF. Установлено, что форма кривой, характеризующей возбудимость ткани в зависимости от интенсивности и длительности действия раздражителя, однотипна для самых разнообразных тканей. Различия между ними касаются только абсолютного значения соответствующих величин и, прежде всего, времени, т. е. возбудимые ткани отличаются друг от друга временной константой раздражения. Лабильность можно измерить, раздражая ткань электрическим током различной частоты. Момент, когда ткань произойдёт преобразование ритма (ткань перестанет воспроизводить заданный ритм без изменений) и будет лабильностью данной ткани. Единицы её измерения - количество воспроизводимых импульсов за единицу времени [имп./сек.(мин.), и т. д. ]. Проводимость можно охарактеризовать расстоянием, преодолённым импульсом за единицу времени, то есть скоростью распространения импульса.
История вопроса.
Изучался этот вопрос давно. Многих учёных интересовал тот факт, что некоторые животные способны генерировать <электрический ток. Это явление было названо <животное электричество. К этому времени относится начало исследований итальянского физиолога и врача Л. Гальвани, заложивших основу учения о Б. п. Многолетний научный спор (1791-97) между Л. Гальвани и физиком А. Вольта о природе <животного электричества завершился двумя крупными открытиями: были получены факты о существовании биоэлектрических явлений в живых тканях и открыт новый принцип получения электрического тока с помощью разнородных металлов - создан гальванический элемент (вольтов столб). Правильная оценка наблюдений Гальвани стала возможной лишь после применения достаточно чувствительных электроизмерительных приборов - гальванометров. Первые такие исследования были проведены итальянским физиком К. Маттеуччи (1837). Систематическое изучение биопотенциалов было начато немецким физиологом Э. Дюбуа-Реймоном (1848), который доказал существование биопотенциалов в нервах и мышцах в покое и при возбуждении. Но ему не удалось (в силу большой инерционности гальванометра) зарегистрировать быстрые, длящиеся тысячные доли секунды колебания биопотенциалов при проведении импульсов вдоль нервов и мышц. В 1886 немецкий физиолог Ю. Бернштейн проанализировал форму потенциала действия; французский учёный Э. Ж. Марей (1875) применил для записи колебаний потенциалов бьющегося сердца капиллярный электрометр; русский физиолог Н. Е. Введенский использовал (1883) для прослушивания ритмических разрядов импульсов в нерве и мышце телефон, а голландский физиолог В. Эйнтховен (1903) ввёл в эксперимент и клиническую практику струнный гальванометр - высокочувствительный и малоинерционный прибор для регистрации электрических токов в тканях. Значительный вклад в изучение биопотенциалов внесли русские физиологи: В. В. Правдич-Неминский (1913-21) впервые зарегистрировал электроэнцефалограмму, А. Ф. Самойлов (1929) исследовал природу нервно-мышечной передачи возбуждения, а Д. С. Воронцов (1932) открыл следовые колебания биопотенциалов, сопровождающие потенциал действия в нервных волокнах. Дальнейший прогресс в изучении биопотенциалов был тесно связан с успехами электроники, позволившими применить в физиологическом эксперименте электронные усилители и осциллографы (работы американских физиологов Г. Бишопа, Дж. Эрлангера и Г. Гассера в 30-40-х гг. 20в.). Изучение биопотенциалов в отдельных клетках и волокнах стало возможным с разработкой микроэлектродной техники. Важное значение для выяснения механизмов генерации биопотенциалов имело использование гигантских нервных волокон головоногих моллюсков, главным образом кальмара. Диаметр этих волокон в 50 - 100 раз больше, чем у позвоночных животных, он достигает 0,5-1 мм, что позволяет вводить внутрь волокна микроэлектроды, инъецировать в протоплазму различные вещества и т.п. Изучение ионной проницаемости мембраны гигантских нервных волокон позволило английским физиологам А. Ходжкину, А. Хаксли и Б. Катцу (1947-52) сформулировать современную мембранную теорию возбуждения.
Потенциал покоя. Потенциал действия.
Потенциал покоя (ПП) - разность потенциалов между содержимым клетки (волокна) и внеклеточной жидкостью; скачок потенциала локализуется на поверхностной мембране, при этом её, внутренняя сторона заряжена электроотрицательно по отношению к наружной. Потенциал покоя обусловлен неравенством концентраций, ионов Na+, К+ и Cl- по обе стороны клеточной мембраны и неодинаковой её проницаемостью для этих ионов. В нервных и мышечных клетках потенциал покоя участвует в поддержании состояния готовности молекулярной структуры мембраны к возбуждению в ответ на действие раздражителя. Все воздействия на клетку, вызывающие длительное стойкое снижение потенциала покоя. (например, нарушение обмена веществ, повышение внеклеточного содержания ионов К+, действие сильного электрического тока и т.д.), ведут к снижению возбудимости клетки или к полной утрате ею способности к генерации потенциалов действия. У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60-90мв, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной.
Концентрация К+ в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na+ примерно в 100 раз ниже, чем для К+, диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К+. В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl-, диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na+ и К+ падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na+ значительно превышает скорость увеличения проницаемости мембраны для К+. Неравенство концентраций ионов К+ и Na+ (или Ca+) внутри и снаружи клетки (волокна) поддерживается специальным механизмом (т. н. <натриевым насосом), выталкивающим ионы Na+ из клетки и нагнетающим ионы К+ в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ. Работа таких механизмов обеспечивается, как правило, энергией, выделяемой при расщеплении аденозинтрифосфорной кислоты (АТФ); таким образом, ионные насосы одновременно выполняют функцию ферментов, расщепляющих АТФ и называемых АТФ-азами. Активный перенос Na+ из клетки сопряжён с транспортом К+ в обратном направлении и осуществляется особой ферментной системой - транспортной Na, К, - стимулируемой аденозинтрифосфатазой, локализованной в клеточной мембране. Последняя, гидролизуя аденозинтрифосфорную кислоту (АТФ), высвобождает энергию, которая и затрачивается на активный перенос катионов. Работа насоса в целом зависит от уровня метаболизма клетки.
Все живые организмы и любая их клетка обладают раздражимостью, т. е. способностью отвечать на внешнее раздражение изменением обмена веществ.
Наряду с раздражимостью три вида ткани: нервная, мышечная и железистая - обладают возбудимостью. В ответ на раздражение в возбудимых тканях возникает процесс возбуждения.
Возбуждение представляет собой сложную биологическую реакцию. Обязательными признаками возбуждения являются изменение мембранного потенциала, усиление обмена веществ (повышение потребления О 2 , выделение СО 2 и тепла) и возникновение деятельности, присущей данной ткани: мышца сокращается, железа выделяет секрет, нервная клетка генерирует электрические импульсы. В момент возбуждения ткань из состояния физиологического покоя переходит к присущей ей деятельности.
Следовательно, возбудимостью называют способность ткани отвечать на раздражение возбуждением. Возбудимость - это свойство ткани, тогда как возбуждение - это процесс, ответная реакция на раздражение.
Важнейшим признаком распространяющегося возбуждения является возникновение нервного импульса, или потенциала действия, благодаря которому возбуждение не остается на месте, а проводится по возбудимым тканям. Раздражителем, вызывающим возбуждение, может быть любой агент внешней или внутренней среды (электрический, химический, механический, термический и др.) при условии, что он является достаточно сильным, действует достаточно долго и нарастание его силы происходит достаточно быстро.
Биоэлектрические явления
Биоэлектрические явления - "животное электричество" было открыто в 1791 г. итальянским ученым Гальвани. Данные современной мембранной теории происхождения биоэлектрических явлений получены Ходжкиным, Кацом и Хаксли в исследованиях, проведенных с гигантским нервным волокном кальмара (диаметром 1 мм) в 1952 г.
Плазматическая мембрана клетки (плазмолемма), ограничивающая снаружи цитоплазму клетки, имеет
толщину около 10 нм и состоит из двойного слоя липидов, в который погружены глобулы белков (молекулы, свернутые в клубки или спирали). Белки выполняют функции ферментов, рецепторов, транспортных систем, ионных каналов. Они либо частично, либо целиком погружены в липидный слой мембраны (рис. 13). В состав мембраны входит также небольшое количество углеводов.
Рис. 13. Модель клеточной мембраны как жидкой мозаики из липидов и белков - поперечный разрез (Стерки П., 1984). а - липиды; в - белки
Сквозь мембрану движутся различные вещества в клетку и из клетки. Регуляция этого процесса - одна из основных функций мембраны. Основными ее свойствами являются избирательная и изменчивая проницаемость. Для одних веществ она служит барьером, для других - входными воротами. Вещества могут проходить через мембрану по закону концентрационного градиента (диффузия от большей концентрации к меньшей), по электрохимическому градиенту (разная концентрация заряженных ионов), путем активного транспорта - работа натрий-калиевых насосов.
Мембранный потенциал, или потенциал покоя. Между наружной поверхностью клетки и ее цитоплазмой существует разность потенциалов порядка 60 - 90 мВ (милливольт) , называемая мембранным потенциалом, или потенциалом покоя. Его можно обнаружить при помощи микроэлектродной методики. Микроэлектрод представляет собой тончайший стеклянный капилляр с диаметром кончика 0,2 - 0,5 мкм. Его заполняют раствором электролита (КС1). Второй электрод обычных размеров погружают в раствор Рингера, в котором находится исследуемый объект. Через усилитель биопотенциалов электроды подводят к осциллографу. Если под микроскопом с помощью микроманипулятора микроэлектрод ввести внутрь нервной клетки, нервного или мышечного волокна, то в момент прокола осциллограф покажет разность потенциалов - потенциал покоя (рис. 14). Микроэлектрод настолько тонок, что он практически не повреждает мембраны.

Рис. 14. Измерение потенциала покоя мышечного волокна (А) при помощи внутриклеточного микроэлектрода (схема). М - микроэлектрод; И - индифферентный электрод. Луч на экране осциллографа показан стрелкой
Мембрайно-ионная теория объясняет происхождение потенциала покоя неодинаковой концентрацией несущих электрические заряды К + , Na + и Сl - внутри и вне клетки и различной проницаемостью для них мембраны.
В клетке в 30 - 50 раз больше К + и в 8 - 10 раз меньше Na + , чем в тканевой жидкости. Следовательно, внутри клетки преобладают К + , снаружи - Na + . Основным анионом тканевой жидкости является Сl - . В клетке преобладают крупные органические анионы, которые не могут диффундировать сквозь мембрану. (Как известно, катионы имеют положительный заряд, а анионы - отрицательный.) Состояние неодинаковой ионной концентрации по обе стороны плазматической мембраны называют ионной асимметрией. Она поддерживается работой натрий-калиевых насосов, которые непрерывно перекачивают Na + из клетки и К + в клетку. Работа эта осуществляется с затратой энергии, освобождающейся при расщеплении аденозинтрифосфорной кислоты. Ионная асимметрия - физиологическое явление, сохраняющееся пока клетка жива.
В покое проницаемость мембраны значительно выше для К + , чем для Na + . В силу высокой концентрации ионы К + стремятся выйти из клетки наружу. Сквозь мембрану они проникают на наружную поверхность клетки, но дальше уйти не могут. Крупные анионы клетки, для которых мембрана непроницаема, не могут последовать за калием, и скапливаются на внутренней поверхности мембраны, создавая здесь отрицательный заряд, который удерживает электростатической связью проскочившие через мембрану положительно заряженные ионы калия. Таким образом возникает поляризация мембраны, потенциал покоя; по обе ее стороны образуется двойной электрический слой: снаружи из положительно заряженных ионов К + , а внутри из отрицательно заряженных различных крупных анионов.
Потенциал действия. Потенциал покоя сохраняется до тех пор, пока не возникло возбуждение. Под действием раздражителя проницаемость мембраны для Na + повышается. Концентрация Na + снаружи клетки в 10 раз больше, чем внутри нее. Поэтому Na + сначала медленно, а затем лавинообразно устремляются внутрь. Ионы натрия заряжены положительно, поэтому происходит перезарядка мембраны и ее внутренняя поверхность приобретает положительный заряд, а наружная - отрицательный. Таким образом происходит реверсия потенциала, изменение его на обратный знак. Он становится отрицательным снаружи и положительным внутри клетки. Этим объясняется давно известный факт, что возбужденный участок становится электроотрицательным по отношению к находящемуся в покое. Однако повышение проницаемости мембраны для Na + длится недолго; она быстро снижается и повышается для К + . Это вызывает усиление потока положительно заряженных ионов из клетки во внешний раствор. В итоге происходит реполяризация мембраны, ее наружная поверхность приобретает снова положительный заряд, а внутренняя - отрицательный.
Электрические изменения мембраны в процессе возбуждения получили название потенциала действия. Длительность его измеряется тысячными долями секунды (миллисекундами), амплитуда равна 90 - 120 мВ.
Во время возбуждения Na + входят в клетку, а К + выходят наружу. Казалось бы, что концентрация ионов в клетке должна меняться. Как показали опыты, даже многочасовое раздражение нерва и возникновение в нем десятков тысяч импульсов не изменяют содержания в нем Na + и К + . Это объясняется работой натрий-калиевого насоса, который после каждого цикла возбуждения разводит ионы по местам: накачивает К + обратно в клетку и выводит из нее Na + . Насос работает на энергии внутриклеточного обмена веществ. Это доказывается тем, что яды, прекращающие обмен веществ, прекращают работу насоса.
Потенциал действия, возникая в возбужденном участке, становится раздражителем для соседнего невозбужденного участка мышечного или нервного волокна и обеспечивает проведение возбуждения вдоль мышцы или нерва.
Возбудимость различных тканей неодинакова. Наиболее высокой возбудимостью отличаются рецепторы, специализированные структуры, приспособленные к улавливанию изменений во внешней среде и внутренней среде организма. Затем следует нервная, мышечная и железистая ткани.
Мерой возбудимости является порог раздражения, т. е. та наименьшая сила раздражителя, которая способна вызвать возбуждение. Порог раздражения иначе называют реобазой. Чем выше возбудимость ткани, тем меньшей силы раздражитель способен вызвать возбуждение.
Кроме того, возбудимость можно характеризовать тем временем, в течение которого должен действовать раздражитель, чтобы вызвать возбуждение, иначе говоря, порогом времени. Наименьшее время, в течение которого должен действовать электрический ток пороговой силы, чтобы вызвать возбуждение, называется полезным временем. Полезное время характеризует скорость течения процесса возбуждения.
Возбудимость тканей увеличивается в процессе умеренной деятельности и снижается при утомлении. Возбудимость претерпевает фазовые изменения во время возбуждения. Как только в возбудимой ткани возникает процесс возбуждения, она утрачивает способность отвечать на новое, даже сильное раздражение. Это состояние называется абсолютной невозбудимостью, или абсолютной рефрактерной фазой. Через некоторое время возбудимость начинает восстанавливаться. На пороговое раздражение ткань еще не отвечает, но на сильное раздражение отвечает возбуждением, хотя амплитуда возникающего потенциала действия в это время значительно снижена, т. е. процесс возбуждения слаб. Это фаза относительной рефрактерности. После нее возникает фаза повышенной возбудимости или супернормальности. В это время можно вызвать возбуждение очень слабым раздражителем, ниже пороговой силы. Только после этого возбудимость приходит в норму.
Для исследования состояния возбудимости мышечной или нервной ткани наносят два раздражения друг за другом через определенные интервалы. Первое вызывает возбуждение, а второе - тестирующее - испытывает возбудимость. Если на второе раздражение реакции нет, значит, ткань невозбудима; реакция слабая - возбудимость понижена; реакция усилена - возбудимость повышена. Так, если на сердце наносить раздражение во время систолы, то возбуждения не последует, к концу диастолы раздражение вызывает внеочередное сокращение - экстрасистолу, что свидетельствует о восстановлении возбудимости.
На рис. 15 сопоставлены во времени процесс возбуждения, выражением которого служит потенциал действия, и фазовые изменения возбудимости. Видно, что абсолютная рефрактерная фаза соответствует восходящей части пика - деполяризации, фаза относительной рефрактерности - нисходящей части пика - реполяризации мембраны и фаза повышенной возбудимости - отрицательному следовому потенциалу.
Рис. 15. Схемы изменений потенциала действия (а) и возбудимости нервного волокна (б) в различные фазы потенциала действия. 1 - местный процесс; 2 - фаза деполяризации; 3 - фаза реполяризации. Пунктиром на рисунке обозначены потенциал покоя и исходный уровень возбудимости
Похожая информация.
Раздражимость
Раздражимостью называют способность живых систем под влиянием раздражителей переходить из состояния физиологического покоя в состояние активности. Формы проявления раздражимости весьма разнообразны. У одних клеток при этом изменяются форма, структура, их рост и процесс деления; в других образуются различные химические соединения, совершается та или иная работа, движение.
Раздражителем живой клетки или организма как целого может оказаться любое изменение внешней среды или внутреннего состояния организма, если оно достигает определенной величины.
Среди раздражителей различают физические (температура, укол, давление, свет, звук, электрические раздражители), физико-химические (изменения осмотического давления, активной реакции среды, электролитного состава, коллоидного состояния) и химические (химические пищи, химические соединения, образующиеся в организме,- гормоны, продукты обмена веществ и т. п.).
Рис. 17. Первый (I ) и второй (II) опыты
Естественными раздражителями клеток, вызывающими их деятельность, являются нервные импульсы.
Возбудимость
Клетки нервной и мышечной тканей приспособлены к осуществлению быстрых реакций на раздражение. Клетки этих тканей называют возбудимыми, а их способность отвечать на раздражение возбуждением называют возбудимостью.
Возбуждение способно перемещаться из одного места клетки в другое, из одной клетки в другую.
 Рис. 18.
Последовательные этапы приготовления нервно-мышечного препарата.
Рис. 18.
Последовательные этапы приготовления нервно-мышечного препарата.
Возбуждение характеризуется комплексом химических, функциональных, физико-химических, электрических явлений. Обязательным признаком возбуждения является изменение электрического состояния поверхностной клеточной мембраны. Именно электрические явления обеспечивают проведение возбуждения в возбудимых тканях.
Биоэлектрические явления
Возникновение и распространение возбуждения связано с изменением электрического заряда живой ткани, с так называемыми биоэлектрическими явлениями.
Электрические явления у животных известны давно. Еще в 1776 г. они были описаны у электрического ската. Началом же экспериментального изучения электрических явлений в животных тканях следует считать опыты итальянского врача Луиджи (1791). В опытах он использовал препараты задних конечностей лягушки, соединенных с позвоночником. Подвешивая эти препараты на медном крючке к железным перилам балкона, он обратил внимание, что, когда конечности лягушки раскачивались ветром, их мышцы сокращались при каждом прикосновении к перилам. На основании этого пришел к выводу, что подергивания лапок были вызваны «животным электричеством», зарождающимся в спинном мозге лягушки и передаваемым по металлическим проводникам (крючку и перилам балкона) к мышцам препарата.
 Рис. 19.
Схема вторичного сокращения.
Рис. 19.
Схема вторичного сокращения.
Против этого положения Гальвани о «животном электричестве» выступил физик Александр . В 1792 г. повторил опыты Гальвани и установил, что описанные Гальвани явления нельзя считать «животным электричеством». В опыте Гальвани источником тока служил не спинной мозг лягушки, а цепь, образованная из разнородных металлов - меди и железа.
Был прав. Первый опыт Гальвани не доказывал наличия «животного электричества», но эти исследования привлекли внимание ученых к изучению электрических явлений в живых образованиях.
В ответ на возражение Вольта Гальвани произвел второй опыт, уже без участия металлов. При этом также наблюдалось сокращение лапки лягушки.
Опыт Гальвани. Для этого обездвижьте лягушку и перережьте ее поперек в области верхних грудных позвонков. Захватив остаток позвоночника салфеточкой, снимите с задних конечностей кожу, а затем пинцетом удалите остатки внутренностей. Становятся хорошо видны нервные стволики крестцового сплетения, расположенные с обеих сторон позвоночника пучками. Подведите под оба пучка нервных волокон одну пластинку пинцета Гальвани, а другой пластинкой пинцета прикоснитесь к нервам сверху. Мышцы лапок при этом сокращаются (рис. 17, I). Пинцет Гальвани состоит из цинковой и медной пластинок. Объясните, почему сокращаются мышцы лапок в опыте Гальвани.
 Рис. 20.
Рис. 20.
А - одноканальный; Б - двухканальный; справа - кончик электрода около тела нейрона
А теперь приготовьте нервно-мышечный препарат лягушки.
Основные этапы приготовления нервно-мышечного препарата приведены на рисунке 18.
Лягушку обездвижьте. Возьмите ее левой рукой за бедра (в этом положении хорошо выделяется позвоночник) и перережьте позвоночник на 1-1,5 см выше места отхождения тазовых костей (рис. 18, 1 ). Свисающую переднюю часть туловища и внутренности удалите. Остаток позвоночника крепко держите пинцетом или левой рукой. Другим пинцетом захватите кожу около позвоночника и тяните ее вниз, чтобы, выворачивая, снять с конечностей (рис. 18, 2 ). Конечности положите на чистую тарелку и залейте раствором Рингера. Руки вымойте или тщательно вытрите от слизи, покрывающей кожу лягушки. Захватите пинцетом или рукой кусочек позвоночника и подогните его вниз так, чтобы конечности висели под углом к позвоночнику и хорошо выделялась копчиковая кость (рис. 18,3).
Осторожно вырежьте копчиковую кость. Ножницы при этом держите как можно ближе к кости, чтобы не повредить идущие параллельно с обеих сторон нервы. Вырезав копчик, положите препарат на тарелку и разделите его на две половины. Для этого перережьте вдоль сначала остаток позвоночника, а затем лобковое сочленение (рис. 18, 4).
 Рис. 21.
А
с помощью микроэлектрода:
Рис. 21.
А
с помощью микроэлектрода:
М - микроэлектрод; И - индифферентный электрод. Луч на экране осциллографа Б показывает, что до прокола микроэлектро дом мембраны разность потенциалов меж ду М и И была равна нулю. В момент прокола (показан стрелкой) обнаружена разность потенциалов, указывающая, что внутренняя сторона мембраны заряжена электроотрицательно по отношению к ее наружной поверхности.
Одну конечность оставьте как запасную, сохраняя ее в растворе Рингера; другую положите на спинную сторону и отделите ножницами подвздошную кость. Захватив пинцетом кусочек позвоночника, отведите в сторону седалищный нерв и удалите подвздошную кость. При помощи двух пинцетов раздвиньте мышцу на спинной поверхности бедра по средней линии (рис. 18, 5). Осторожно не касаясь ножницами и пинцетом нерва, отделите его от окружающих тканей, вдоль бедра до колен. (Лучше это делать стеклянным крючком.) Нерв отведите в сторону и освободите бедренную кость от мышц (рис. 18, б). На голени отделите от кости икроножную мышцу, подрезав ахиллово сухожилие, и привяжите к нему нитку. Голень и стопу отрежьте ниже колена (рис. 18, 7). Препарат положите в стакан с раствором Рингера.
Проделайте второй опыт Гальвани (сокращение без металла). Для этого нервно-мышечный препарат положите на дощечку. Отрежьте кусочек мышцы и стеклянным крючком быстро набросьте нерв препарата на пораненный участок мышцы так, чтобы он одновременно коснулся поврежденной и неповрежденной поверхности мышцы (рис. 17, II). Мышца при этом сокращается. Объясните, почему это происходит.
Гальвани все же оказался прав в своем утверждении о существовании «животного электричества», что позже было подтверждено исследованиями других ученых.
В этом отношении интересны опыты Маттеучю, получившие название вторичного сокращения.
На мышцу одного нервно-мышечного препарата набросьте нерв другого нервно-мышечного препарата (рис. 19) и раздражайте электрическим током нерв первого препарата. Вы наблюдаете сокращение мышцы и второго препарата. Это объясняется тем, что при возбуждении в мышце первого препарата возникают токи действия, которые вызывают возбуждение второго нервно-мышечного препарата.
В дальнейшем в изучение биоэлектрических явлений очень важный вклад внесли русские ученые, среди них И. М. Сеченов, обнаруживший с помощью гальванометра электрические явления в головном мозге, Н. Е. Введенский, А. Ф. Самойлов и др.
В настоящее время имеются весьма совершенные, высокочувствительные приборы (электронно-лучевые трубки с электронными усилителями), позволяющие регистрировать электрические явления в тканях и органах.
Потенциал покоя и потенциал действия мышц
Между наружной поверхностью клетки и ее цитоплазмой в состоянии покоя существует разность потенциалов около 60-90 мВ, причем поверхность клетки заряжена электроположительно по отношению к цитоплазме. Эту разность потенциалов называют потенциалом покоя или мембранным потенциалом. Точное измерение потенциала покоя возможно только с помощью микроэлектродов, предназначенных для внутриклеточного отведения токов, очень мощных усилителей и чувствительных регистрирующих приборов - осциллографов.
 Рис. 22.
Потенциал действия аксона кальмара, отводимый с помощью внутриклеточного электрода.
Рис. 22.
Потенциал действия аксона кальмара, отводимый с помощью внутриклеточного электрода.
Микроэлектрод (рис. 20) представляет собой тонкий стеклянный капилляр, кончик которого имеет диаметр около микрона. Этот капилляр заполняют солевым раствором, погружают в него металлический электрод и соединяют с усилителем и осциллографом (рис. 21). Как только микроэлектрод прокалывает покрывающую клетку мембрану, луч осциллографа отклоняется вниз из своего исходного положения и устанавливается на новом уровне. Это свидетельствует о наличии разности потенциалов между наружной и внутренней поверхностью клеточной мембраны.
Наиболее полно происхождение потенциала покоя объясняет так называемая мембран но-ионная теория. Согласно этой теории все клетки покрыты мембраной, имеющей неодинаковую проницаемость для различных ионов. В связи с этим внутри клетки в цитоплазме в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем на поверхности клетки. В состоянии покоя клеточная мембрана более проницаема для ионов калия, чем для ионов натрия.
Диффузия положительно заряженных ионов калия из цитоплазмы на поверхность клетки придает наружной поверхности мембраны положительный заряд.
Таким образом, поверхность клетки в покое несет на себе положительный заряд, тогда как внутренняя сторона мембраны оказывается заряженной отрицательно за счет ионов хлора, аминокислот и других крупных органических анионов, которые через мембрану практически не проникают.
Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя, в этом участке возникает возбуждение, проявляющееся в быстром колебании мембранного потенциала и называемое потенциалом действия.
Потенциал действия можно зарегистрировать с помощью электродов, приложенных к внешней поверхности волокна (внеклеточное отведение), либо микроэлектрода, введенного в цитоплазму (внутриклеточное отведение) (рис. 22).
При введении микроэлектрода в нервное волокно сначала регистрируется потенциал величиной около 60 мВ. Это мембранный потенциал.
Потенциал действия представляет собой резкий скачок мембранного потенциала. При этом происходит не только изменение отрицательного потенциала покоя от величины - 60 мВ до нуля, но и переход за нулевую линию на несколько десятков милливольт, так что амплитуда потенциала действия превышает потенциал покоя и составляет приблизительно 90 мВ.
В потенциале действия различают его пик (спайк, по терминологии английских авторов) и следовые потенциалы. Во время пика мембранный потенциал меняет свой знак.
Причина возникновения потенциала действия - изменение ионной проницаемости мембраны. При раздражении проницаемость клеточной мембраны для ионов натрия повышается. стремится внутрь клетки, так как, во-первых, он заряжен положительно и его влекут внутрь электростатические силы, вовторых, концентрация его внутри клетки невелика. В покое клеточная мембрана была малопроницаемой для этого иона. Раздражение изменило проницаемость мембраны, и поток положительно заряженных ионов натрия из внешней среды клетки в цитоплазму значительно превысил поток ионов калия из клетки наружу. В результате произошло извращение потенциала мембраны (фаза деполяризации). Внутренняя поверхность мембраны стала заряженной положительно, а наружная вследствие потери положительно заряженных ионов натрия - отрицательно. В этот момент и регистрируется пик потенциала действия.
Повышение проницаемости мембраны для ионов натрия продолжается очень короткое время. Вслед за этим в клетке возникают восстановительные процессы, приводящие к тому, что проницаемость мембраны для ионов натрия вновь понижается, а для ионов калия возрастает. Поскольку ионы калия также заряжены положительно, выходя из клетки, они восстанавливают исходные отношения снаружи и внутри клетки (фаза реполяризации).
Накопления натрия внутри клетки при многократном возбуждении ее не происходит, потому что эвакуируется из нее постоянно за счет действия специального биохимического механизма, называемого «натриевым насосом». Есть данные и об активном транспорте калия с помощью «натрий-калиевого насоса».
Таким образом, согласно мембранной теории в происхождении биоэлектрических явлений решающее значение имеет избирательная проницаемость клеточной мембраны, обусловливающая разный ионный состав на поверхности и внутри клетки, а следовательно, и разный заряд этих поверхностей. Следует заметить, что многие положения мембранной теории все еще дискуссионны и нуждаются в углубленной разработке.
Итак, биоэлектрические свойства клеток, обусловленные неравномерным распределением минеральных ионов, играют ведущую роль в процессах клеточного возбуждения. Возникновение потенциала действия является наиболее характерным показателем возбуждения в клетках, тканях и органах. Поэтому электрофизиологические исследования широко применяются в экспериментальных лабораториях и в клиниках.
Значение регистрации биоэлектрических явлений. Регистрация электрических потенциалов в живом организме дает возможность судить об изменениях многих физиологических функций. Так как электрическая активность многих органов человека в норме имеет типичные и постоянные характеристики, методы электрофизиологии широко используются для диагностики болезней в практической медицине. Большие успехи достигнуты в тонком распознавании болезней сердца, нервной системы, мышц. Электрофизиологические методы сыграли важную роль в решении многих проблем космической физиологии. С помощью методов телеметрии оказалось возможным передавать информацию о состоянии сердечной мышцы, деятельности мозга, скелетной мускулатуры и других органов в условиях невесомости, перегрузок.
Изучение электрической активности центральной нервной системы легло в основу конструирования сложных счетно-решающих электронных машин.
Под электроэнцефалографией понимают запись биоэлектрических явлений, протекающих в головном мозге, преимущественно в коре больших полушарий головного мозга.
Для отведения биотоков от различных структур головного мозга используют различные конструкции электродов. В эксперименте на животных электроды можно ввести через кости черепа прямо в нужный участок головного мозга. Такие «вживленные» электроды долго удерживаются в мозгу специальным креплением и позволяют изучать электрическую активность определенных участков головного мозга при различных состояниях животного.
 Рис. 23.
Рис. 23.
В настоящее время возможна длительная регистрация электрических явлений даже в отдельных клетках мозга с помощью микроэлектродов.
При записи биотоков мозга человека - электроэнцефалограммы - пользуются обычно серебряными электродами, имеющими вид пластинки размером с двухкопеечную монету. Электроды на голове испытуемого человека укрепляются с помощью шлемов-сеток. Шлемы изготовляют из эластичных резиновых тяжей, натяжение которых регулируют. Шлемы, плотно прилегая к голове испытуемого, надежно удерживают электроды (рис. 23).
Запись биотоков мозга производится на приборах - электроэнцефалографах, имеющих разную конструкцию и включающих несколько усилителей биотоков, осциллографы и сложный пульт управления ими.
В настоящее время выпускают приборы, позволяющие регистрировать одновременно электрическую активность от 2 до 32 точек мозга и более.
Для точного анализа электрических явлений, возникающих в головном мозге и других органах, используют электронно-вычислительные машины.
Электрическая активность мозга человека носит ритмический характер. Электроды, расположенные на поверхности головы, дают возможность зарегистрировать биотоки сразу от многих клеток мозга, лежащих под ними. Поэтому общий характер энцефалограммы оказывается очень сложным. Вместе с тем удалось установить, что наиболее выраженных и часто встречающихся ритмов колебания электрической активности немного. Названия этих ритмов условно обозначены греческими буквами α, β, δ, ϑ (рис. 25, табл. 4).
Таблица 4
Ритмы электроэцефалограммы
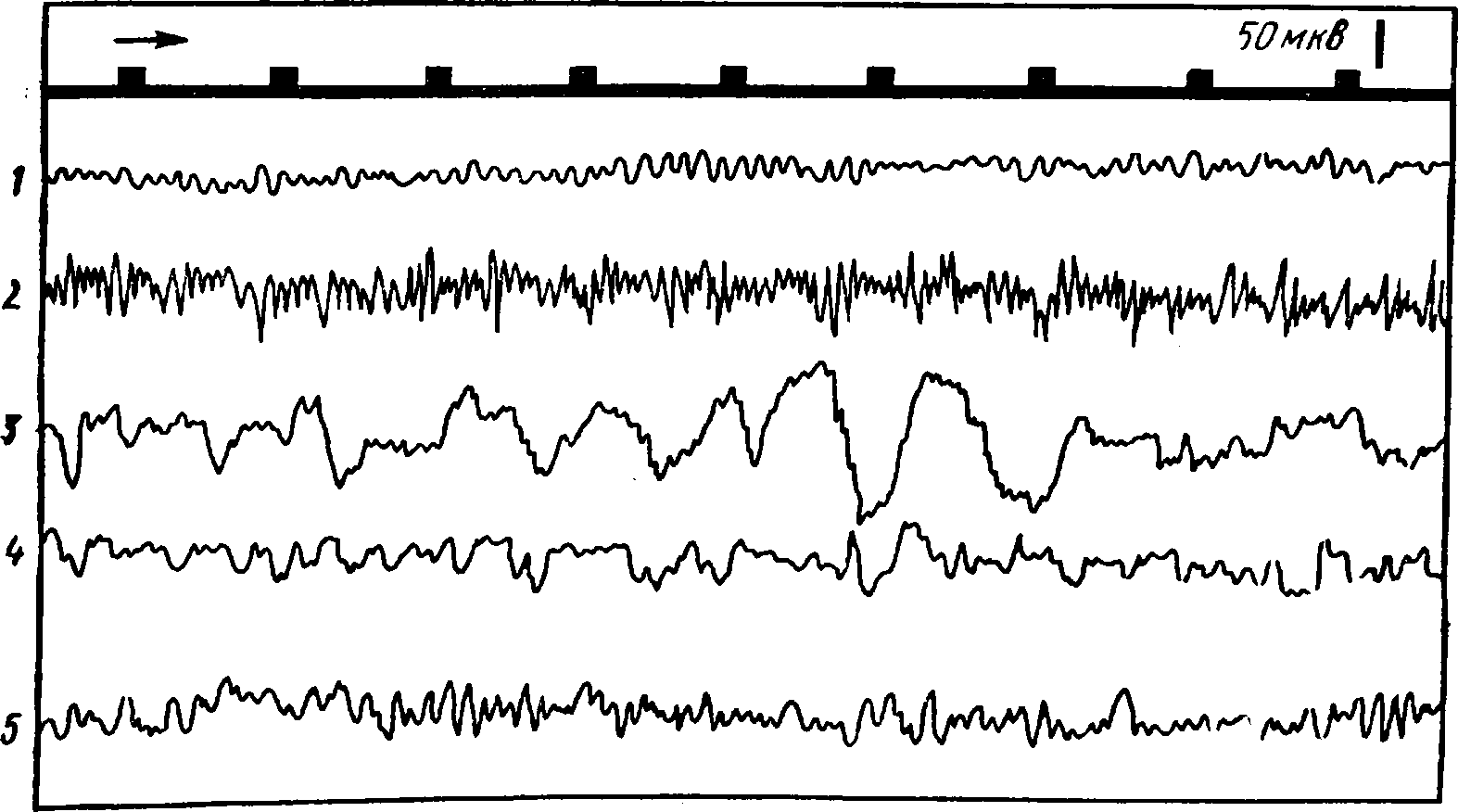 Рис. 25.
Рис. 25.
1 -альфа-ритм- 2 - бета-ритм; 3 - дельта-ритм; 4- тета-ритм; 5 -веретена; вверх у - отметка времени
При ограниченном поступлении центростремительных импульсов в исследуемый участок мозга обычно наблюдаются медленные волны с большим размахом колебаний. Если в кору поступают многочисленные импульсы, то клетки в этом участке могут находиться в разных стадиях возбуждения и общая электрическая активность над этим участком характеризуется частыми колебаниями с небольшой амплитудой типа бета-ритма. На рисунке 26 приведена электроэнцефалограмма с затылочной области коры (там располагается зрительная зона коры больших полушарий). На ней отчетливо виден переход альфа-ритма в бета-ритм и обратно при открывании и закрывании глаз, т. е. при увеличении и снижении потока центростремительных импульсов к зрительной зоне коры.
Современная техника позволяет регистрировать энцефалограмму у человека на расстоянии и даже в условиях космического полета.
Получил достаточно широкое распространение и метод исследования электрической активности мышц - электромиография.
 Для отведения биопотенциалов мышц человека используют накожные металлические электроды диаметром 10 мм. Электроды укрепляют на исследуемой мышце эластической манжетой; между ними и кожей находится обычно специальна
я паста, улучшающая контакт с телом и электропроводность. Колебания биопотенциалов мышц имеют также ритмический характер, только частота их и амплитуда значительно больше, чем при записи электроэнцефалограммы. Усиление мышечной активности сопровождается увеличением амплитуды и колебаний электромиограммы. При утомлении мышц частота колебаний, как правило, падает.
Для отведения биопотенциалов мышц человека используют накожные металлические электроды диаметром 10 мм. Электроды укрепляют на исследуемой мышце эластической манжетой; между ними и кожей находится обычно специальна
я паста, улучшающая контакт с телом и электропроводность. Колебания биопотенциалов мышц имеют также ритмический характер, только частота их и амплитуда значительно больше, чем при записи электроэнцефалограммы. Усиление мышечной активности сопровождается увеличением амплитуды и колебаний электромиограммы. При утомлении мышц частота колебаний, как правило, падает.
Рис. 26. Изменения электроэнцефалограммы затылочной области коры, показывающие переход от альфа-ритма к бета-ритму при открывании глаз (стрелка вверх) и восстановление альфа-ритма при закрывании глаз (стрелка вниз).
На рисунке 27 представлены электромиограммы трехглавой и двуглавой мышц плеча при работе напильником. На них отчетливо видно усиление колебаний при возбуждении мышц. Электромиография позволяет установить степень участия тех или иных мышц в выполняемом движении. Особенно важен этот метод для установления наличия паралича мышц при некоторых заболеваниях. Так, с помощью электромиографии можно обнаружить паралич дыхательных мышц при полиомиелите до того, как остановится . А это очень важно для принятия необходимых мер (перевод на управляемое с помощью специальной аппаратуры) с целью спасения жизни человеку.
 Рис. 27.
Электромиограммы трехглавой (1)
и двуглавой (2)
мышц
плеча у рабочего при опиловке.
Рис. 27.
Электромиограммы трехглавой (1)
и двуглавой (2)
мышц
плеча у рабочего при опиловке.
Данные электромиографии используются при создании активных протезов и управляемых манипуляторов.
«Искусственная рука» - одно из первых устройств, управляемых с помощью биотоков мышц. Это активный протез, воспроизводящий нормальные движения кисти. Такой активный протез имеет браслет для отведения биотоков мышц сохранившейся культи. На браслете укреплены металлические чашечки, заполненные токопроводящей пастой. Отводимые с помощью браслета мышечные токи усиливаются и поступают в специальный блок на протезе, где формируется управляющий сигнал. Сигнал приводит в действие миниатюрный двигатель, который обеспечивает движение протеза. Сам протез выполняется из пластических материалов, сходных по форме, размерам и цвету с нормальной рукой.
Передача возбуждения в синапсах
Возбуждение от одной нервной клетки к другой или от нервного волокна к мышечной или железистой клетке передается с помощью синап сов.
Аксоны большинства ней ронов, подходя к другим нервным клеткам, ветвятся и образуют многочисленные окончания на телах этих кле ток и их дендринах (рис. 28). Такие места контактов называют синапсами.
 Рис. 28.
Расположение синапсов на теле нейрона и на его дендритах (по Хаачу и Барру)
Рис. 28.
Расположение синапсов на теле нейрона и на его дендритах (по Хаачу и Барру)
Количество синапсов на теле одного нейрона достигает 100 и больше, а на дендритах одного нейрона - нескольких тысяч. Одно нервное волокно может образовать до 10 000 синапсов на многих нервных клетках.
Синапс имеет сложное строение (рис. 29). Он образован двумя мембранами - пресинаптической и постси наптической, между которыми есть синоптическая щель. Пресинаптическая мембрана находится на нервном окончании. Нервные окончания в центральной нервной системе имеют вид пуговок, колечек или бляшек. На теле пирамидной клетки насчитывается несколько тысяч нервных окончаний. Каждая синаптическая пуговка покрыта пресинаптической мембраной. Постсинаптическая мембрана находится на теле или дендритах нейрона, к которому передается нервный импульс. В пресинаптической области обычно наблюдаются большие скопления митохондрий.
Возбуждение через синапсы передается химическим путем с помощью особого — посредника, или медиатора, находящегося в синаптической бляшке. В синаптической бляшке медиатор находится в синаптических пузырьках, каждый из которых содержит до 2000 молекул медиатора. В разных синапсах вырабатываются разные медиаторы. Чаще всего в качестве медиатора служит ацетилхолин, адреналин или норадреналин в некоторых синапсах - глютаминовая кислота.
Каков же механизм передачи возбуждения в синапсах? Приход нервного импульса в пресинаптическое окончание сопровождается синхронным выбросом в синаптическую щель медиатора из синаптических пузырьков, расположенных в непосредственной близости от нее. Размеры этой щели очень малы (около 200 Å), и медиатор, быстро достигая постсинаптической мембраны, взаимодействует с ее веществом. В результате этого взаимодействия структура постсинаптической мембраны временно изменяется, проницаемость ее для ионов натрия повышается, что приводит к перемещению ионов и, как следствие, появлению возбуждающего постсинаптического потенциала. Когда этот потенциал достигает определенной величины, возникает распространяющее возбуждение - потенциал действия.
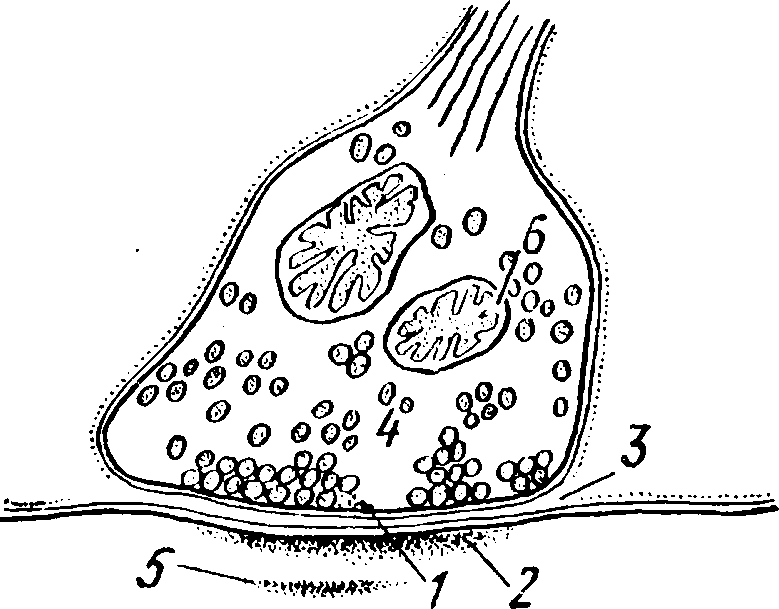 Рис. 29.
Рис. 29.
1 - пресинаптическая мембрана; 2 - постсинаптическая мембрана; 3 - синаптичес кая щель; 4 - синаптические пузырьки; 5 - нейропротофибриллы; 6 - митохондрии.
Через очень короткое время (несколько миллисекунд) медиаторы разрушаются специальными ферментами.
В настоящее время подавляющее большинство нейрофизиологов признает существование в спинном мозге и в различных отделах головного мозга двух качественно различных типов синапсов - возбуждающих и тормозящих. Полагают, что в специализированных тормозящих нейронах, в нервных окончаниях аксонов вырабатывается особый медиатор, оказывающий тормозящее воздействие на последующий нейрон. Природа этого медиатора еще окончательно не установлена. В коре больших полушарий таким медиатором некоторые авторы считают гамма-амино-масляную кислоту.
Под влиянием приходящего по аксону тормозящего нейрона импульса в синаптическую щель выделяется медиатор, который вызывает специфические изменения в постсинаптической мембране. Суть этих изменений состоит в том, что медиатор торможения, взаимодействуя с веществом постсинаптической мембраны, увеличивает ее проницаемость для ионов калия и хлора. Внутри клетки относительно увеличивается число анионов. В результате происходит не снижение величины внутреннего заряда мембраны (как это имеет место в возбуждающем синапсе), а повышение внутреннего заряда постсинаптической мембраны. Это ведет к возникновению Главная Анатомия человека
Токи покоя Наличие разности потенциалов можно обнаружить, если к поврежденной мышце приложить электроды осциллографа таким образом, чтобы один из них... История изучения человека Анатомия физиология человека Место человека в природе Клетка Обмен веществ клетки Деление... Обмен веществ Регуляция деятельности организма Нервная система Рефлекс и рефлекторная дуга Безусловные и условные рефлексы Кровь Плазма крови Реакция крови...










